


–õ—é–Ī–ł–ľ –ľ–Ķ—ā–ĺ–ī –Ĺ–į –≤—Ā–ł—á–ļ–ł –ļ—Ä–Ķ–į—Ü–ł–ĺ–Ĺ–ł—Ā—ā–ł –Ķ –ī–į –≤–Ĺ—É—ą–į–≤–į—ā, —á–Ķ –Ķ–≤–ĺ–Ľ—é—Ü–ł–ĺ–Ĺ–ł—Ā—ā–ł—ā–Ķ –ł–ľ–į—ā –į–Ī—Ā—É—Ä–ī–Ĺ–ł –ł–ī–Ķ–ł, –ļ–ĺ–ł—ā–ĺ —ā–Ķ —Ā–į–ľ–ł—Ź —Ā–ł –ł–∑–ľ–ł—Ā–Ľ—Ź—ā. –ē—ā–ĺ —Ü–ł—ā–į—ā –ĺ—ā –ļ–Ĺ–ł–≥–į—ā–į –Ĺ–į –•–į—Ä—É–Ĺ –Į—Ö–į "–ē–≤–ĺ–Ľ—é—Ü–ł–ĺ–Ĺ–Ĺ–į—ā–į –ł–∑–ľ–į–ľ–į":
"–Ď–Ķ–∑–≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—ā–Ķ –ł–ľ–į—ā —ā–≤—ä—Ä–ī–į —á–į—Ā—ā, –Ĺ–į–ľ–ł—Ä–į—Č–į —Ā–Ķ –ĺ—ā –≤—ä–Ĺ—ą–Ĺ–į—ā–į –ł–ľ —Ā—ā—Ä–į–Ĺ–į, –ī–ĺ–ļ–į—ā–ĺ —Ä–ł–Ī–ł—ā–Ķ —Ā–į –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł –ł —ā–≤—ä—Ä–ī–į—ā–į –ł–ľ —á–į—Ā—ā –Ķ —Ä–į–∑–Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ–į –≤—ä–≤ –≤—ä—ā—Ä–Ķ—ą–Ĺ–ĺ—Ā—ā—ā–į –Ĺ–į —ā—Ź–Ľ–ĺ—ā–ĺ –ł–ľ. –ó–į –ĺ—Ā—ä—Č–Ķ—Ā—ā–≤—Ź–≤–į–Ĺ–Ķ—ā–ĺ –Ĺ–į –Ķ–ī–Ĺ–į —ā–į–ļ–į–≤–į –≤–Ķ–Ľ–ł—á–Ķ—Ā—ā–≤–Ķ–Ĺ–į ‚Äú–Ķ–≤–ĺ–Ľ—é—Ü–ł—Ź‚ÄĚ –Ķ –Ĺ–Ķ–ĺ–Ī—Ö–ĺ–ī–ł–ľ–ĺ –Ņ—Ä–ł–ļ–Ľ—é—á–≤–į–Ĺ–Ķ—ā–ĺ –Ĺ–į –ľ–ł–Ľ–ł–į—Ä–ī–ł —Ā—ā–į–ī–ł–ł, –Ķ—ā–ĺ –∑–į—Č–ĺ –Ī–ł —ā—Ä—Ź–Ī–≤–į–Ľ–ĺ –ī–į —Ā–į –ĺ—Ā—ā–į–Ĺ–į–Ľ–ł –≤–ļ–į–ľ–Ķ–Ĺ–Ķ–Ľ–ĺ—Ā—ā–ł –Ĺ–į –ľ–ł–Ľ–ł–į—Ä–ī–ł –Ĺ–į –Ī—Ä–ĺ–Ļ –ľ–Ķ–∂–ī–ł–Ĺ–Ĺ–ł —Ą–ĺ—Ä–ľ–ł, –ĺ—ā—Ä–į–∑—Ź–≤–į—Č–ł —ā–Ķ–∑–ł —Ā—ā–į–ī–ł–ł." - —ā—É–ļ
–ö–į–ļ –ĺ—ā –Ī—Ä—ä–ľ–Ī–į—Ä —Č–Ķ —Ā—ā–į–Ĺ–Ķ —Ä–ł–Ī–į? –ú–Ĺ–ĺ–≥–ĺ –Ņ—Ä–ĺ—Ā—ā–ĺ - —ā—Ä—Ź–Ī–≤–į –ī–į –≥–ĺ –ĺ–Ī—ä—Ä–Ĺ–Ķ–ľ –Ĺ–į–ĺ–Ņ–į–ļ–ł! –Ę–ĺ–≤–į –Ķ —ą–Ķ–≥–į –Ķ—Ā—ā–Ķ—Ā—ā–≤–Ķ–Ĺ–ĺ, –Ĺ–ĺ...
–ě–Ī—ä—Ä–Ĺ–į—ā–ł –Ĺ–į–ĺ–Ņ–į–ļ–ł
–°–Ņ–ĺ—Ä–Ķ–ī —ā–ĺ–≤–į, –ļ–į–ļ–≤–ĺ —Ā–Ķ —Ā–Ľ—É—á–≤–į —Ā –Ī–Ľ–į—Ā—ā–ĺ–Ņ–ĺ—Ä–į—ā–į (–Ņ—ä—Ä–≤–ł—á–Ĺ–ł—Ź –ĺ—ā–≤–ĺ—Ä, –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ –ĺ—ā –≤–≥—ä–≤–į–Ĺ–Ķ—ā–ĺ –Ĺ–į –∑–į—Ä–ĺ–ī–ł—ą–į) –ľ–Ĺ–ĺ–≥–ĺ–ļ–Ľ–Ķ—ā—ä—á–Ĺ–ł—ā–Ķ –Ī–ł–Ľ–į—ā–Ķ—Ä–į–Ľ–Ĺ–ł (–ī–≤—É—Ā—ā—Ä–į–Ĺ–Ĺ–ĺ —Ā–ł–ľ–Ķ—ā—Ä–ł—á–Ĺ–ł) –∂–ł–≤–ĺ—ā–Ĺ–ł —Ā–Ķ –ī–Ķ–Ľ—Ź—ā –Ĺ–į –ī–≤–Ķ –≥—Ä—É–Ņ–ł. –Ę–Ķ–∑–ł, –Ņ—Ä–ł –ļ–ĺ–ł—ā–ĺ –Ī–Ľ–į—Ā—ā–ĺ–Ņ–ĺ—Ä–į—ā–į —Ā—ā–į–≤–į —É—Ā—ā–į, —Ā–Ķ –Ĺ–į—Ä–ł—á–į—ā –Ņ—ä—Ä–≤–ł—á–Ĺ–ĺ—É—Ā—ā–Ĺ–ł (Prostomia). –Ę–į–ļ–ł–≤–į —Ā–į –Ņ–ĺ–≤–Ķ—á–Ķ—ā–ĺ –Ī–Ķ–∑–≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł, –≤ —ā.—á. —á–Ľ–Ķ–Ĺ–Ķ—Ā—ā–ĺ–Ĺ–ĺ–≥–ł—ā–Ķ —Ā —Ö–ł—ā–ł–Ĺ–ĺ–≤–į –ĺ–Ī–≤–ł–≤–ļ–į, –∑–į –ļ–ĺ–ł—ā–ĺ –Ĺ–į–ľ–Ķ–ļ–≤–į –•.–Į.
|
¬† –Ē—Ä—É–≥–į—ā–į –≤—ä–∑–ľ–ĺ–∂–Ĺ–ĺ—Ā—ā –Ķ –Ī–Ľ–į—Ā—ā–ĺ–Ņ–ĺ—Ä–į—ā–į –ī–į —Ā—ā–į–Ĺ–Ķ –į–Ĺ—É—Ā. –£—Ā—ā–į—ā–į –Ņ—Ä–ł —ā—Ź—Ö —Ā–Ķ –ĺ–Ī—Ä–į–∑—É–≤–į –ĺ—ā –≤—ā–ĺ—Ä–ł—á–Ĺ–ĺ –≤–ī–Ľ—ä–Ī–≤–į–Ĺ–Ķ –Ĺ–į –Ņ—Ä–ĺ—ā–ł–≤–ĺ–Ņ–ĺ–Ľ–ĺ–∂–Ĺ–į—ā–į —Ā—ā—Ä–į–Ĺ–į –Ĺ–į –Ī–Ľ–į—Ā—ā–ĺ–Ņ–ĺ—Ä–į—ā–į. –Ę–ĺ–≤–į —Ā—ā–į–≤–į –Ņ—Ä–ł –≤—Ā–ł—á–ļ–ł —Ö–ĺ—Ä–ī–ĺ–≤–ł (–≤ —ā.—á. –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł) –ł –ľ–į–Ľ—ä–ļ –Ī—Ä–ĺ–Ļ –Ī–Ķ–∑–≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł, –Ĺ–į–Ņ—Ä–ł–ľ–Ķ—Ä –Ī–ĺ–ī–Ľ–ĺ–ļ–ĺ–∂–ł—ā–Ķ. –ě–Ī—Č–ĺ —ā–Ķ–∑–ł –∂–ł–≤–ĺ—ā–Ĺ–ł —Ā–Ķ –Ĺ–į—Ä–ł—á–į—ā –≤—ā–ĺ—Ä–ł—á–Ĺ–ĺ—É—Ā—ā–Ĺ–ł (Deuterostomia) - —Ā–Ņ—Ä–į–≤–ļ–į –ĺ—ā —Ā–į–Ļ—ā–į –Ĺ–į –ú–į–Ļ—Ź –ú–į—Ä–ļ–ĺ–≤–į. –° –ī—Ä—É–≥–ł –ī—É–ľ–ł –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—ā–Ķ –ł —á–Ľ–Ķ–Ĺ–Ķ—Ā—ā–ĺ–Ĺ–ĺ–≥–ł—ā–Ķ —Ā–į —Ā–Ķ —Ä–į–∑–ī–Ķ–Ľ–ł–Ľ–ł –ĺ—Č–Ķ –≤ –Ĺ–į—á–į–Ľ–ĺ—ā–ĺ –Ĺ–į –Ķ–≤–ĺ–Ľ—é—Ü–ł—Ź—ā–į –Ĺ–į –ľ–Ĺ–ĺ–≥–ĺ–ļ–Ľ–Ķ—ā—ä—á–Ĺ–ł—ā–Ķ, –ĺ—Č–Ķ –Ņ—Ä–Ķ–ī–ł –ö–į–ľ–Ī—Ä–ł–Ļ—Ā–ļ–ł—Ź –≤–∑—Ä–ł–≤. –ė —Ā–į –ĺ—ā –ī–≤–Ķ —Ä–į–∑–Ľ–ł—á–Ĺ–ł –Ĺ–į–ī–≥—Ä—É–Ņ–ł, —ā–ĺ–Ľ–ļ–ĺ–≤–į —Ä–į–∑–Ľ–ł—á–Ĺ–ł, —ā–į —á–į–ļ –ĺ–Ī—ä—Ä–Ĺ–į—ā–ł –Ĺ–į–ī–ĺ–Ľ—É —Ā –≥–Ľ–į–≤–į—ā–į! –Ě–į —Ā—Ö–Ķ–ľ–į—ā–į: a - –≥–į—Ā—ā—Ä—É–Ľ–į; b - –Ņ—ä—Ä–≤–ł—á–Ĺ–ĺ—É—Ā—ā–Ĺ–ł (Prostomia); c - –≤—ā–ĺ—Ä–ł—á–Ĺ–ĺ—É—Ā—ā–Ĺ–ł (Deuterostomia); A - –≤—Ö–ĺ–ī (—É—Ā—ā–į), Anterior; P - –ł–∑—Ö–ĺ–ī (–į–Ĺ—É—Ā) Posterior |
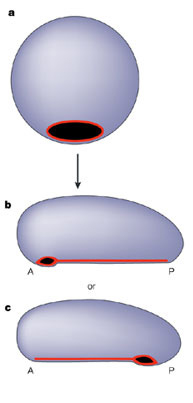 —Ā—Ö–Ķ–ľ–į: Different strategies for midline formation in bilaterians —Ā—Ö–Ķ–ľ–į: Different strategies for midline formation in bilaterians |
 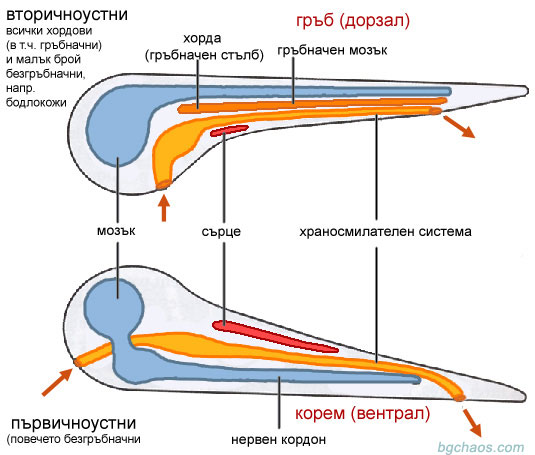
| –ě—Ä–ł–≥–ł–Ĺ–į–Ľ–Ĺ–į —Ā—Ö–Ķ–ľ–į: Etienne Geoffroy Saint-Hilaire revu et corrig√© (–Ņ—Ä–Ķ—Ä–į–Ī–ĺ—ā–Ķ–Ĺ–į) |
–ü—Ä–Ķ–∑ 1822–≥ —Ą—Ä–Ķ–Ĺ—Ā–ļ–ł—Ź—ā –∑–ĺ–ĺ–Ľ–ĺ–≥ –Ė–ĺ—Ą—Ä–ĺ–į –°–Ķ–Ĺ—ā-–ė–Ľ–Ķ—Ä (Geoffroy Saint-Hilaire) –Ņ—É–Ī–Ľ–ł–ļ—É–≤–į —Ā—ā–į—ā–ł—Ź, –ļ—ä–ī–Ķ—ā–ĺ –ĺ—ā–Ī–Ķ–Ľ—Ź–∑–į–Ľ, —á–Ķ –≤—ä—ā—Ä–Ķ—ą–Ĺ–ł—ā–Ķ —Ā–ł—Ā—ā–Ķ–ľ–ł –Ņ—Ä–ł –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—ā–Ķ —Ā–į —Ä–į–∑–Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ–ł –ĺ–≥–Ľ–Ķ–ī–į–Ľ–Ĺ–ĺ –Ĺ–į –Ņ–ĺ–ī–ĺ–Ī–Ĺ–ł—ā–Ķ –ł–ľ –Ņ—Ä–ł –Ī–Ķ–∑–≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—ā–Ķ. –Ě–į–Ņ—Ä–ł–ľ–Ķ—Ä –Ņ—Ä–ł –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—ā–Ķ —Ü–Ķ–Ĺ—ā—Ä–į–Ľ–Ĺ–į—ā–į –Ĺ–Ķ—Ä–≤–Ĺ–į —Ā–ł—Ā—ā–Ķ–ľ–į —Ā–Ķ –Ĺ–į–ľ–ł—Ä–į –ĺ—ā–ļ—ä–ľ –≥—ä—Ä–Ī–į (–ī–ĺ—Ä–∑–į–Ľ–Ĺ–ĺ), —Ā—ä—Ä—Ü–Ķ—ā–ĺ –Ķ –ĺ—ā–ļ—ä–ľ –ļ–ĺ—Ä–Ķ–ľ–Ĺ–į—ā–į —Ā—ā—Ä–į–Ĺ–į (–≤–Ķ–Ĺ—ā—Ä–į–Ľ–Ĺ–ĺ), –į –Ņ—Ä–ł —á–Ľ–Ķ–Ĺ–Ķ—Ā—ā–ĺ–Ĺ–ĺ–≥–ł—ā–Ķ –Ķ –ĺ–Ī—Ä–į—ā–Ĺ–ĺ. –ß–Ķ—Ä–Ĺ–ł—Ź—ā –ī—Ä–ĺ–Ī —Ā–Ķ –Ĺ–į–ľ–ł—Ä–į –ĺ—ā –Ľ—Ź–≤–į—ā–į —Ā—ā—Ä–į–Ĺ–į –Ĺ–į —Ā—ā–ĺ–ľ–į—Ö–į –Ĺ–į —á–Ľ–Ķ–Ĺ–Ķ—Ā—ā–ĺ–Ĺ–ĺ–≥–ł—ā–Ķ –ł –ĺ—ā–ī—Ź—Ā–Ĺ–ĺ –Ņ—Ä–ł –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—ā–Ķ –∂–ł–≤–ĺ—ā–Ĺ–ł, –Ņ–ĺ—Ā–ĺ–ļ–į—ā–į –Ĺ–į –ļ—Ä—ä–≤–Ĺ–ł—Ź –Ņ–ĺ—ā–ĺ–ļ –≤ —á–Ľ–Ķ–Ĺ–Ķ—Ā—ā–ĺ–Ĺ–ĺ–≥–ł –ł –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—ā–Ķ —Ā—ä—Č–ĺ –Ķ –Ņ—Ä–ĺ—ā–ł–≤–ĺ–Ņ–ĺ–Ľ–ĺ–∂–Ĺ–į –ł —ā–į–ļ–į –Ĺ–į—ā–į—ā—ä–ļ.
–Ē–į¬†–Ņ—Ä–ĺ–ī—ä–Ľ–∂–ł–ľ –ĺ—Č–Ķ –Ņ–ĺ-–Ĺ–į—ā–į—ā—ä–ļ¬†- –ź–ļ–ĺ –ĺ–Ī—ä—Ä–Ĺ–Ķ–ľ –Ĺ–į–ĺ–Ņ–į–ļ–ł –ļ–į—ā–ĺ —Ä—ä–ļ–į–≤–ł—Ü–į —ā—Ź–Ľ–ĺ—ā–ĺ –Ĺ–į –Ķ–ī–Ĺ–ĺ –Ī–Ķ–∑–≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ĺ —Ā –Ķ–ļ–∑–ĺ—Ā–ļ–Ķ–Ľ–Ķ—ā, –ĺ—Ä–≥–į–Ĺ–ł—ā–Ķ —Ā—ä—Č–ĺ –Ī–ł—Ö–į —Ā–Ķ –ĺ–Ī—ä—Ä–Ĺ–į–Ľ–ł –ĺ–≥–Ľ–Ķ–ī–į–Ľ–Ĺ–ĺ, –į —Ā–ļ–Ķ–Ľ–Ķ—ā—ä—ā –Ī–ł –ĺ—Ā—ā–į–Ĺ–į–Ľ –≤—ä—ā—Ä–Ķ. –Ē–į–Ľ–ł –Ĺ–Ķ –Ī–ł –ľ–ĺ–≥–Ľ–ĺ –ī–į —Ā—ā–į–Ĺ–Ķ –Ľ–Ķ—Ā–Ĺ–ĺ —Ā –Ķ–ī–Ĺ–į "–ľ–į–ļ—Ä–ĺ–ľ—É—ā–į—Ü–ł—Ź"?
–°–ľ–Ķ–Ľ–į—ā–į –∑–į –≤—Ä–Ķ–ľ–Ķ—ā–ĺ —Ā–ł —Ö–ł–Ņ–ĺ—ā–Ķ–∑–į –Ĺ–į –Ė–ĺ—Ą—Ä–ĺ–į –°–Ķ–Ĺ—ā-–ė–Ľ–Ķ—Ä –Ī–ł–Ľ–į, —á–Ķ –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—ā–Ķ –ł –Ī–Ķ–∑–≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—ā–Ķ –∂–ł–≤–ĺ—ā–Ĺ–ł –ł–ľ–į—ā –Ķ–ī–ł–Ĺ –ł —Ā—ä—Č –Ņ–Ľ–į–Ĺ –∑–į —ā—Ź–Ľ–ĺ—ā–ĺ, –Ĺ–ĺ –≤ –Ĺ—Ź–ļ–į–ļ—ä–≤ –ľ–ĺ–ľ–Ķ–Ĺ—ā –≤ —Ā–≤–ĺ—Ź—ā–į –Ķ–≤–ĺ–Ľ—é—Ü–ł—Ź —Ā–į –ĺ–Ī—ä—Ä–Ĺ–į—ā–ł –Ĺ–į–ĺ–Ņ–į–ļ–ł! –°–Ľ–Ķ–ī –ĺ—ā–ļ—Ä–ł—ā–ł—Ź—ā–į –≤ –≥–Ķ–Ĺ–Ķ—ā–ł–ļ–į—ā–į, —ā–į–∑–ł –ł–ī–Ķ—Ź—ā–į –Ĺ–Ķ –ł–∑–≥–Ľ–Ķ–∂–ī–į —ā–ĺ–Ľ–ļ–ĺ–≤–į "–Ľ—É–ī–į".
–ó–į –ī–į —Ā–Ķ –Ņ–ĺ—Ź–≤–ł –ĺ–ļ–ĺ –Ĺ–į –≥–Ľ–į–≤–į—ā–į –Ĺ–į –ļ–į–ļ—ä–≤—ā–ĺ –ł –ī–į –Ķ –ĺ—Ä–≥–į–Ĺ–ł–∑—ä–ľ, –Ī–ł–Ľ —ā–ĺ–Ļ –≥—Ä—ä–Ī–Ĺ–į—á–Ķ–Ĺ –ł–Ľ–ł –Ĺ–Ķ, –Ķ –Ĺ–Ķ–ĺ–Ī—Ö–ĺ–ī–ł–ľ —Ā–į–ľ–ĺ –Ķ–ī–ł–Ĺ –≥–Ķ–Ĺ: Pax-6. –ė –Ĺ–Ķ —Ā–į–ľ–ĺ –Ņ–ĺ—Ā–Ľ–Ķ–ī–ĺ–≤–į—ā–Ķ–Ľ–Ĺ–ĺ—Ā—ā—ā–į –Ĺ–į —ā–ĺ–∑–ł –≥–Ķ–Ĺ –Ķ –ł–ī–Ķ–Ĺ—ā–ł—á–Ĺ–į –Ņ—Ä–ł –ľ–ł—ą–ļ–ł –ł –ľ—É—Ö–ł—ā–Ķ –ī—Ä–ĺ–∑–ĺ—Ą–ł–Ľ–į, –Ĺ–ĺ –ł —Ä–į–∑–ľ—Ź–Ĺ–į—ā–į –Ĺ–į —ā–ĺ–∑–ł –≥–Ķ–Ĺ –Ĺ–Ķ –Ņ—Ä–ł—á–ł–Ĺ—Ź–≤–į –∑–Ĺ–į—á–ł—ā–Ķ–Ľ–Ĺ–į –Ņ—Ä–ĺ–ľ—Ź–Ĺ–į. –ď–Ķ–Ĺ—ä—ā Pax-6 –ĺ—ā –ľ–ł—ą–ļ–į, –Ņ—Ä–Ķ–Ĺ–Ķ—Ā–Ķ–Ĺ –≤ –ī—Ä–ĺ–∑–ĺ—Ą–ł–Ľ–į –Ņ—Ä–Ķ–ī–ł–∑–≤–ł–ļ–≤–į –ĺ–Ī—Ä–į–∑—É–≤–į–Ĺ–Ķ—ā–ĺ –Ĺ–į –ĺ–ļ–ĺ –Ĺ–į –ī—Ä–ĺ–∑–ĺ—Ą–ł–Ľ–į, –ļ–į–ļ—ā–ĺ –ł –ĺ–Ī—Ä–į—ā–Ĺ–ĺ—ā–ĺ. –Ę—Ä–į–ī–ł—Ü–ł–ĺ–Ĺ–Ĺ–ł—ā–Ķ –≤—ä–∑–≥–Ľ–Ķ–ī–ł –∑–į "–Ĺ–Ķ–∑–į–≤–ł—Ā–ł–ľ –≤—ä–Ĺ—ą–Ķ–Ĺ –≤–ł–ī" –Ĺ–į –ĺ—á–ł—ā–Ķ (–ł –ī—Ä—É–≥–ł –Ķ–Ľ–Ķ–ľ–Ķ–Ĺ—ā–ł –ĺ—ā –Ņ–Ľ–į–Ĺ–į –ł –ĺ—Ä–≥–į–Ĺ–ł–∑–į—Ü–ł—Ź—ā–į) –≤ —Ä–į–∑–Ľ–ł—á–Ĺ–ł—ā–Ķ –≤–ł–ī–ĺ–≤–Ķ —Ā–Ķ –ĺ–ļ–į–∑–≤–į –Ĺ–į–Ņ—ä–Ľ–Ĺ–ĺ –Ņ–ĺ–≥—Ä–Ķ—ą–Ĺ–į.
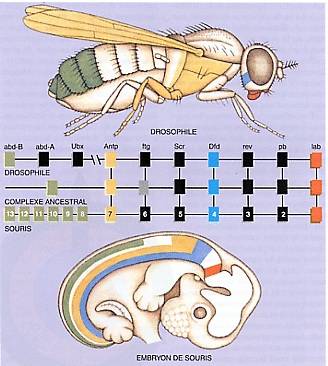
–ü—Ä–ł —Ā—Ä–į–≤–Ĺ–Ķ–Ĺ–ł–Ķ –Ĺ–į –Ņ—Ä–ĺ—ā–Ķ–ł–Ĺ–ĺ–≤–ł—ā–Ķ –Ņ–ĺ—Ā–Ľ–Ķ–ī–ĺ–≤–į—ā–Ķ–Ľ–Ĺ–ĺ—Ā—ā–ł —Ā–Ķ –ĺ–ļ–į–∑–≤–į, —á–Ķ homeobox –≥–Ķ–Ĺ–ł—ā–Ķ, –ļ–ĺ–ł—ā–ĺ —É—á–į—Ā—ā–≤–į—ā –≤ —Ä–Ķ–≥—É–Ľ–ł—Ä–į–Ĺ–Ķ—ā–ĺ –Ĺ–į –ľ–ĺ–ī–Ķ–Ľ–ł—ā–Ķ –Ĺ–į –į–Ĺ–į—ā–ĺ–ľ–ł—á–Ĺ–ĺ—ā–ĺ —Ä–į–∑–≤–ł—ā–ł–Ķ (–ľ–ĺ—Ä—Ą–ĺ–≥–Ķ–Ĺ–Ķ–∑–į—ā–į) –≤ –∂–ł–≤–ĺ—ā–Ĺ–ł, –≥—ä–Ī–ł –ł —Ä–į—Ā—ā–Ķ–Ĺ–ł—Ź —Ā–į –ł–ī–Ķ–Ĺ—ā–ł—á–Ĺ–ł –≤ –∂–ł–≤–ĺ—ā–ł–Ĺ—Ā–ļ–ĺ—ā–ĺ —Ü–į—Ä—Ā—ā–≤–ĺ. –ė–Ľ—é—Ā—ā—Ä–į—Ü–ł—Ź: L'origine des innovations √©volutives dans le monde animal |

–ď–Ķ–Ĺ—ä—ā, –ļ–ĺ–Ļ—ā–ĺ –Ņ—Ä–Ķ–ī–ł–∑–≤–ł–ļ–≤–į –ĺ–Ī—Ä–į–∑—É–≤–į–Ĺ–Ķ—ā–ĺ –Ĺ–į –Ĺ–Ķ–≤—Ä–į–Ľ–Ĺ–į—ā–į —ā—Ä—ä–Ī–į (—Ā—ä–ĺ—ā–≤–Ķ—ā—Ā—ā–≤–į—Č–į –Ĺ–į –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—Ź –ľ–ĺ–∑—ä–ļ –Ņ—Ä–ł –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł –∂–ł–≤–ĺ—ā–Ĺ–ł) –Ņ—Ä–ł –ī—Ä–ĺ–∑–ĺ—Ą–ł–Ľ–į—ā–į –Ķ –≥–Ķ–Ĺ SOG, –į –Ņ—Ä–ł –ļ—Ä–į—Ā—ā–į–≤–į—ā–į –∂–į–Ī–į Xenopus (–ļ–į–ļ—ā–ĺ –Ķ –≤—ä–≤ –≤—Ā–ł—á–ļ–ł –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł –∂–ł–≤–ĺ—ā–Ĺ–ł) –Ķ –ī—Ä—É–≥ –≥–Ķ–Ĺ, –Ĺ–į—Ä–Ķ—á–Ķ–Ĺ chordin. –ě—Č–Ķ –≤–Ķ–ī–Ĺ—ä–∂ —Ā–Ķ –Ņ–ĺ—ā–≤—ä—Ä–∂–ī–į–≤–į —Ö–ł–Ņ–ĺ—ā–Ķ–∑–į—ā–į –Ĺ–į –°–Ķ–Ĺ—ā-–ė–Ľ–Ķ—Ä. –Ě–Ķ —Ā–į–ľ–ĺ –Ņ–ĺ—Ā–Ľ–Ķ–ī–ĺ–≤–į—ā–Ķ–Ľ–Ĺ–ĺ—Ā—ā—ā–į –Ĺ–į —ā–Ķ–∑–ł –ī–≤–į –≥–Ķ–Ĺ–į –Ķ –Ņ–ĺ—á—ā–ł –Ķ–ī–Ĺ–į–ļ–≤–į, –Ĺ–ĺ –ł –∑–į–ľ—Ź–Ĺ–į—ā–į –ł–ľ –Ķ–ī–ł–Ĺ —Ā –ī—Ä—É–≥ –Ĺ–Ķ –≤–Ľ–ł—Ź–Ķ –≤—ä—Ä—Ö—É –ļ—Ä–į–Ļ–Ĺ–ĺ—ā–ĺ —ā—Ź–Ľ–ĺ. –°–į–ľ–ĺ —á–Ķ —Ā–į –ĺ–Ī—ä—Ä–Ĺ–į—ā–ł –ĺ–≥–Ľ–Ķ–ī–į–Ľ–Ĺ–ĺ. –ė–Ľ—é—Ā—ā—Ä–į—Ü–ł—Ź: Des chim√®res, des clones et des g√®nes |
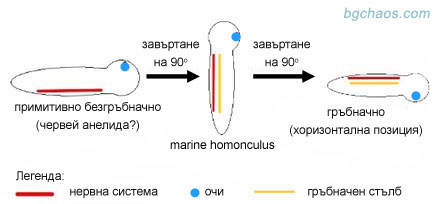
–ė–Ĺ—ā–Ķ—Ä–Ķ—Ā–Ĺ–į —Ö–ł–Ņ–ĺ—ā–Ķ–∑–į –ł–ľ–į —Ą—Ä–Ķ–Ĺ—Ā–ļ–ł—Ź –ł—Ö—ā–ł–ĺ–Ľ–ĺ–≥ –§—Ä–į–Ĺ—Ā–ĺ–į –ī—Ć–ĺ –°–į—Ä (Fran√ßois de Sarre), –ļ–ĺ–Ļ—ā–ĺ –ĺ–Ī—Ź—Ā–Ĺ—Ź–≤–į –Ņ—Ä–Ķ—Ö–ĺ–ī–į –ĺ—ā –Ī–Ķ–∑–≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł –ļ—ä–ľ –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł —Ā –ī–≤–Ķ –∑–į–≤—ä—Ä—ā–į–Ĺ–ł—Ź –Ĺ–į 90–ĺ .
–°–≤—ä—Ä–∑–≤–į—Č–ĺ –∑–≤–Ķ–Ĺ–ĺ –Ķ –Ķ–ī–Ĺ–ĺ –≤–Ķ—Ä—ā–ł–ļ–į–Ľ–Ĺ–ĺ —Ā—ā–ĺ—Ź—Č–ĺ —Ā—ā—Ä–į–Ĺ–Ĺ–ĺ —Ā—ä—Č–Ķ—Ā—ā–≤–ĺ, –Ĺ–į—Ä–Ķ—á–Ķ–Ĺ–ĺ –ľ–ĺ—Ä—Ā–ļ–ł —Ö–ĺ–ľ—É–Ĺ–ļ—É–Ľ—É—Ā (marine homonculus), –ļ–ĺ–Ķ—ā–ĺ –Ķ –Ņ–Ľ—É–≤–į–Ľ–ĺ –≤–Ķ—Ä—ā–ł–ļ–į–Ľ–Ĺ–ĺ –Ņ–ĺ –ī—Ä–Ķ–≤–Ĺ–ł—ā–Ķ –ľ–ĺ—Ä–Ķ—ā–ĺ —Ā –ĺ–≥—Ä–ĺ–ľ–Ĺ–į—ā–į —Ā–ł –≥–Ľ–į–≤–į —ą–į–ľ–į–Ĺ–ī—É—Ä–į. –°–Ņ–ĺ—Ä–Ķ–ī –Ĺ–Ķ–≥–ĺ –≤—Ā–ł—á–ļ–ł –ĺ—Ā—ā–į–Ĺ–į–Ľ–ł –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł —Ā–į —Ā–Ķ —Ä–į–∑–≤–ł–Ľ–ł –ĺ—ā –Ĺ–Ķ–≥–ĺ —á—Ä–Ķ–∑ "–ī–Ķ—Ö—É–ľ–į–Ĺ–ł–∑–į—Ü–ł—Ź", –į —á–ĺ–≤–Ķ–ļ—ä—ā –ĺ—Ā—ā–į–≤–į –Ĺ–į–Ļ-—Ā–Ľ–į–Ī–ĺ –Ķ–≤–ĺ–Ľ—é–ł—Ä–į–Ľ –≤ —ā–į–∑–ł –ļ–ĺ–ľ–Ņ–į–Ĺ–ł—Ź.
–Ę–ĺ–≤–į: Aquatic proto-people and the theory hypothesis of initial bipedalism –Ķ —Ā—ā–į—ā–ł—Ź –Ĺ–į –į–Ĺ–≥–Ľ–ł–Ļ—Ā–ļ–ł –Ņ–ĺ —ā–Ķ–ľ–į—ā–į
 |
| –ě—Ä–ł–≥–ł–Ĺ–į–Ľ–Ĺ–į —Ā—Ö–Ķ–ľ–į: Etienne Geoffroy Saint-Hilaire revu et corrig√© (–Ņ—Ä–Ķ—Ä–į–Ī–ĺ—ā–Ķ–Ĺ–į) |
 
–Ě—É–∂–Ĺ–ĺ –Ľ–ł –Ķ –ī–į —Ā–Ņ–ĺ–ľ–Ķ–Ĺ–į–≤–į–ľ, —á–Ķ —ā–Ķ–ĺ—Ä–ł—Ź—ā–į –ľ—É —Ā–Ķ –Ņ—Ä–ł–Ķ–ľ–į –≤ –Ĺ–į—É—á–Ĺ–ł—ā–Ķ —Ā—Ä–Ķ–ī–ł –ļ–į—ā–ĺ –Ņ–ĺ-—Ā–ļ–ĺ—Ä–ĺ –Ķ–ļ–∑–ĺ—ā–ł—á–Ĺ–į, –ĺ—ā–ļ–ĺ–Ľ–ļ–ĺ—ā–ĺ –ł–ľ–į—Č–į —Ā–Ķ—Ä–ł–ĺ–∑–Ĺ–ł –ĺ—Ā–Ĺ–ĺ–≤–į–Ĺ–ł—Ź. –Ē–į –ļ–į–∂–Ķ–ľ, —á–Ķ –Ė–ĺ—Ą—Ä–ĺ–į –°–Ķ–Ĺ—ā-–ė–Ľ–Ķ—Ä –ł –§—Ä–į–Ĺ—Ā–ĺ–į –ī–Ķ –°–į—Ä –Ĺ–Ķ –ĺ—ā–ł–≤–į—ā –ī–ĺ—Ā—ā–į—ā—ä—á–Ĺ–ĺ –ī–į–Ľ–Ķ—á –≤ –ľ–ł–Ĺ–į–Ľ–ĺ—ā–ĺ, –į –ī–ĺ—Ā—ā–į –≤—Ā—ā—Ä–į–Ĺ–ł –≤ —ā—ä—Ä—Ā–Ķ–Ĺ–Ķ –Ĺ–į —Ö–ł–Ņ–ĺ—ā–Ķ—ā–ł—á–Ĺ–ł—Ź —Ä–ĺ–ī–ĺ–Ĺ–į—á–į–Ľ–Ĺ–ł–ļ –Ĺ–į –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—ā–Ķ. –ß–Ķ—Ä–≤–Ķ–ł—ā–Ķ-–į–Ĺ–Ķ–Ľ–ł–ī–ł —Ā–į —ā–≤—ä—Ä–ī–Ķ —Ā–Ņ–Ķ—Ü–ł–į–Ľ–ł–∑–ł—Ä–į–Ĺ–ł - –≤—Ā–ł—á–ļ–ĺ –Ķ —Ā—ā–į–Ĺ–į–Ľ–ĺ –ľ–Ĺ–ĺ–≥–ĺ, –ľ–Ĺ–ĺ–≥–ĺ –Ņ–ĺ-—Ä–į–Ĺ–ĺ.
–í—ā–ĺ—Ä–ł—á–Ĺ–ĺ—É—Ā—ā–Ĺ–ł—ā–Ķ –Ī—Ä–į—ā–ĺ–≤—á–Ķ–ī–ł
–ú–Ķ–∂–ī—É –≤–Ņ—Ä–ĺ—á–Ķ–ľ –ł–ľ–į–ľ–Ķ –Ķ–ī–ł–Ĺ –Ņ—Ä–į–≤–ĺ—Ā—ā–ĺ—Ź—Č –ī–į–Ľ–Ķ—á–Ķ–Ĺ –Ī—Ä–į—ā–ĺ–≤—á–Ķ–ī - –ź—Ā—Ü–ł–ī–ł—Ź—ā–į. –í–Ľ—Ź–≤–ĺ –Ķ –≤—ä–∑—Ä–į—Ā—ā–Ķ–Ĺ –Ķ–ļ–∑–Ķ–ľ–Ņ–Ľ—Ź—Ä, –į –≤–ī—Ź—Ā–Ĺ–ĺ - –ł–∑–Ĺ–Ķ–Ĺ–į–ī–į - –Ľ–į—Ä–≤–į—ā–į –Ĺ–į –į—Ā—Ü–ł–ī–ł—Ź—ā–į, –ļ–ĺ—Ź—ā–ĺ –≤—ä–Ĺ—ą–Ĺ–ĺ –Ĺ–į–Ņ–ĺ–ī–ĺ–Ī—Ź–≤–į –Ņ–ĺ–Ņ–ĺ–≤–į –Ľ—ä–∂–ł—á–ļ–į, –Ĺ–ĺ –Ķ –ľ–Ĺ–ĺ–≥–ĺ –Ņ–ĺ-–Ņ—Ä–ĺ—Ā—ā–ĺ —É—Ā—ā—Ä–ĺ–Ķ–Ĺ–į, –Ĺ–ĺ –ł–ľ–į —Ö–ĺ—Ä–ī–į!
–Ě–Ķ–ļ–į –≤–ł –∑–į–Ņ–ĺ–∑–Ĺ–į—Ź —Ā –ī—Ä—É–≥–ł –Ĺ–į—ą–ł –ī–į–Ľ–Ķ—á–Ĺ–ł –Ī—Ä–į—ā–ĺ–≤—á–Ķ–ī–ł, —Ā—ä—Č–ĺ –ļ–į—ā–ĺ –Ĺ–į—Ā –≤—ā–ĺ—Ä–ł—á–Ĺ–ĺ—É—Ā—ā–Ĺ–ł, –Ĺ–ĺ –Ņ—Ä–Ķ–ī–Ņ–ĺ—á–Ķ–Ľ–ł —Ā–Ņ–ĺ–ļ–ĺ–Ļ—Ā—ā–≤–ł–Ķ—ā–ĺ –Ĺ–į –ľ–ĺ—Ä—Ā–ļ–ĺ—ā–ĺ –ī—ä–Ĺ–ĺ (–Ņ–ĺ–Ĺ—Ź–ļ–ĺ–≥–į –ī–ĺ—Ä–ł —Ā–į –Ņ—Ä–ł–ļ—Ä–Ķ–Ņ–Ķ–Ĺ–ł –Ĺ–Ķ–Ņ–ĺ–ī–≤–ł–∂–Ĺ–ĺ –∑–į –Ĺ–Ķ–≥–ĺ –ļ–į—ā–ĺ –Ņ–ĺ–Ľ–ł–Ņ–ł), –Ņ—Ä–Ķ–ī –Ī–Ľ–Ķ—Ā—ā—Ź—Č–į—ā–į –ļ–į—Ä–ł–Ķ—Ä–į –Ĺ–į –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł;) –ó–į–Ņ–ĺ—á–≤–į–ľ —Ā –Ņ–ĺ-–ī–į–Ľ–Ķ—á–Ĺ–ł—ā–Ķ: –ė–≥–Ľ–ĺ–ļ–ĺ–∂–ł (–Ď–ĺ–ī–Ľ–ĺ–ļ–ĺ–∂–ł) (Echinodermata)
 –ú–ĺ—Ä—Ā–ļ–į –∑–≤–Ķ–∑–ī–į (Asteroidea) –°–Ĺ–ł–ľ–ļ–į: wikidot –ú–ĺ—Ä—Ā–ļ–į –∑–≤–Ķ–∑–ī–į (Asteroidea) –°–Ĺ–ł–ľ–ļ–į: wikidot |
 –ú–ĺ—Ä—Ā–ļ–į –Ľ–ł–Ľ–ł—Ź (Crinoidea) –°–Ĺ–ł–ľ–ļ–į: bio1152 –ú–ĺ—Ä—Ā–ļ–į –Ľ–ł–Ľ–ł—Ź (Crinoidea) –°–Ĺ–ł–ľ–ļ–į: bio1152 |
 –ú–ĺ—Ä—Ā–ļ–ł —ā–į—Ä–į–Ľ–Ķ–∂ (Echinoidea) –°–Ĺ–ł–ľ–ļ–į: wikipedia –ú–ĺ—Ä—Ā–ļ–ł —ā–į—Ä–į–Ľ–Ķ–∂ (Echinoidea) –°–Ĺ–ł–ľ–ļ–į: wikipedia |
 –õ–į—Ä–≤–į –Ĺ–į –ľ–ĺ—Ä—Ā–ļ–į –∑–≤–Ķ–∑–ī–į Asterias rubens –°–Ĺ–ł–ľ–ļ–į: Mic-UK –õ–į—Ä–≤–į –Ĺ–į –ľ–ĺ—Ä—Ā–ļ–į –∑–≤–Ķ–∑–ī–į Asterias rubens –°–Ĺ–ł–ľ–ļ–į: Mic-UK |
 –ú–ĺ—Ä—Ā–ļ–į –ļ—Ä–į—Ā—ā–į–≤–ł—Ü–į (Peniagone diaphana) –°–Ĺ–ł–ľ–ļ–į: MAR-ECO –ú–ĺ—Ä—Ā–ļ–į –ļ—Ä–į—Ā—ā–į–≤–ł—Ü–į (Peniagone diaphana) –°–Ĺ–ł–ľ–ļ–į: MAR-ECO |
 –ú–ĺ—Ä—Ā–ļ–į –ļ—Ä–į—Ā—ā–į–≤–ł—Ü–į (Peniagone diaphana) –°–Ĺ–ł–ľ–ļ–į: MAR-ECO –ú–ĺ—Ä—Ā–ļ–į –ļ—Ä–į—Ā—ā–į–≤–ł—Ü–į (Peniagone diaphana) –°–Ĺ–ł–ľ–ļ–į: MAR-ECO |
 –ú–ĺ—Ä—Ā–ļ–į –ļ—Ä–į—Ā—ā–į–≤–ł—Ü–į Holothuroidea –°–Ĺ–ł–ľ–ļ–į: Awesome wildlife: –ú–ĺ—Ä—Ā–ļ–į –ļ—Ä–į—Ā—ā–į–≤–ł—Ü–į Holothuroidea –°–Ĺ–ł–ľ–ļ–į: Awesome wildlife: |
 –•–ĺ–Ľ–ĺ—ā—É—Ä–ł—Ź –°–Ĺ–ł–ľ–ļ–į: MAR-ECO –•–ĺ–Ľ–ĺ—ā—É—Ä–ł—Ź –°–Ĺ–ł–ľ–ļ–į: MAR-ECO |
–ü–ĺ–Ľ—É—Ö–ĺ—Ä–ī–ĺ–≤–ł (Hemichordata)
 –ú–ĺ—Ä—Ā–ļ–ł —á–Ķ—Ä–≤–Ķ–Ļ Bathypelagic Ctenophore –°–Ĺ–ł–ľ–ļ–į: MAR-ECO –ú–ĺ—Ä—Ā–ļ–ł —á–Ķ—Ä–≤–Ķ–Ļ Bathypelagic Ctenophore –°–Ĺ–ł–ľ–ļ–į: MAR-ECO |
 –ú–ĺ—Ä—Ā–ļ–ł —á–Ķ—Ä–≤–Ķ–Ļ (Enteropneust) –°–Ĺ–ł–ľ–ļ–į: MAR-ECO –ú–ĺ—Ä—Ā–ļ–ł —á–Ķ—Ä–≤–Ķ–Ļ (Enteropneust) –°–Ĺ–ł–ľ–ļ–į: MAR-ECO |
 Phoronida Nemertea –°–Ĺ–ł–ľ–ļ–į: starfish.ch Phoronida Nemertea –°–Ĺ–ł–ľ–ļ–į: starfish.ch |
 Enteropneust –°–Ĺ–ł–ľ–ļ–į: MAR-ECO Enteropneust –°–Ĺ–ł–ľ–ļ–į: MAR-ECO |
–õ–į–Ĺ—Ü–Ķ—ā–Ĺ–ł–ļ (Branchiostoma lanceolatum)
 Branchiostoma belcheri –°–Ĺ–ł–ľ–ļ–į: Xiamen's Branchiostoma belcheri –°–Ĺ–ł–ľ–ļ–į: Xiamen's |
 –°–Ĺ–ł–ľ–ļ–į: Tayn√° Leoncio –°–Ĺ–ł–ľ–ļ–į: Tayn√° Leoncio |
 –°–Ĺ–ł–ľ–ļ–į: Arthur Anker EOL Rapid Response Team –°–Ĺ–ł–ľ–ļ–į: Arthur Anker EOL Rapid Response Team |
 –°–Ĺ–ł–ľ–ļ–į: Dr. Alvaro Migotto –°–Ĺ–ł–ľ–ļ–į: Dr. Alvaro Migotto |
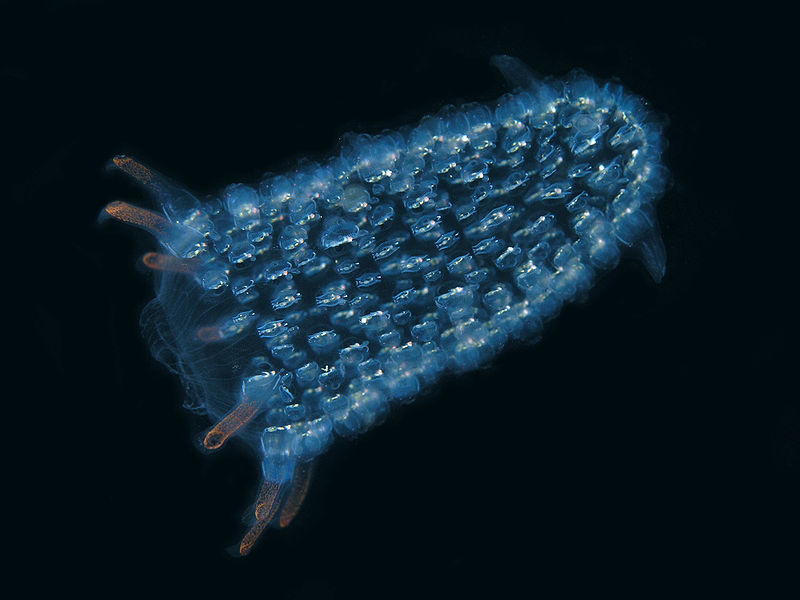
–¶–ł–Ņ–Ķ—Ā—ā–ł (Tunicata)
 Salpa cylindrica solitary –°–Ĺ–ł–ľ–ļ–į: Agrouniversidad Salpa cylindrica solitary –°–Ĺ–ł–ľ–ļ–į: Agrouniversidad  Rhopalaea crassa –°–Ĺ–ł–ľ–ļ–į: Cytochrome Rhopalaea crassa –°–Ĺ–ł–ľ–ļ–į: Cytochrome  Asc√≠dia –°–Ĺ–ł–ľ–ļ–į: El Mundo Oce√°nico Asc√≠dia –°–Ĺ–ł–ľ–ļ–į: El Mundo Oce√°nico |
 –ź–Ņ–Ķ–Ĺ–ī–ł–ļ—É–Ľ–į—Ä–ł—Ź Appendicularia Oikopleura –°–Ĺ–ł–ľ–ļ–į: nature.com –ź–Ņ–Ķ–Ĺ–ī–ł–ļ—É–Ľ–į—Ä–ł—Ź Appendicularia Oikopleura –°–Ĺ–ł–ľ–ļ–į: nature.com  Corella parallelogramma –°–Ĺ–ł–ľ–ļ–į: marinbi.com Corella parallelogramma –°–Ĺ–ł–ľ–ļ–į: marinbi.com |
 –ü–ł—Ä–ĺ—Ā–ĺ–ľ–į (Pyrosoma) –°–Ĺ–ł–ľ–ļ–į: Nick Hobgood wikipedia –ü–ł—Ä–ĺ—Ā–ĺ–ľ–į (Pyrosoma) –°–Ĺ–ł–ľ–ļ–į: Nick Hobgood wikipedia 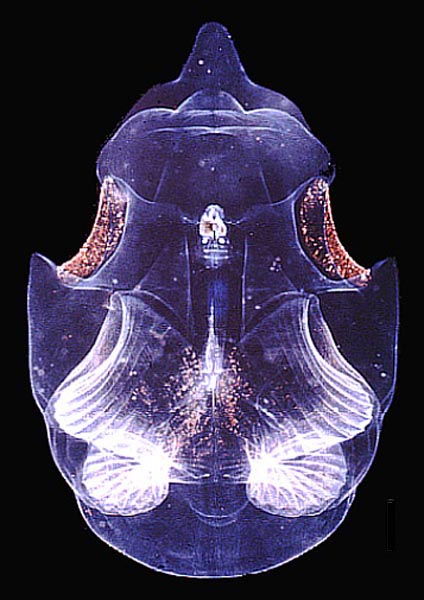 –ě–Ļ–ļ–ĺ–Ņ–Ľ–Ķ–Ļ—Ä–į (Oikopleura labradoriensis) –°–Ĺ–ł–ľ–ļ–į: tieba.baidu.com –ě–Ļ–ļ–ĺ–Ņ–Ľ–Ķ–Ļ—Ä–į (Oikopleura labradoriensis) –°–Ĺ–ł–ľ–ļ–į: tieba.baidu.com |
 Appendicularia Larvacea –°–Ĺ–ł–ľ–ļ–į: wikipedia Appendicularia Larvacea –°–Ĺ–ł–ľ–ļ–į: wikipedia  Clavellina lepadiformis –°–Ĺ–ł–ľ–ļ–į: marinbi.com Clavellina lepadiformis –°–Ĺ–ł–ľ–ļ–į: marinbi.com  Synoicum pulmonaria –°–Ĺ–ł–ľ–ļ–į: marinbi.com Synoicum pulmonaria –°–Ĺ–ł–ľ–ļ–į: marinbi.com |
–Ď–Ķ–∑—á–Ķ–Ľ—é—Ā—ā–Ĺ–ł (Agnatha)
 Hagfish (Myxinidae) –°–Ĺ–ł–ľ–ļ–į: wikitravel.org Hagfish (Myxinidae) –°–Ĺ–ł–ľ–ļ–į: wikitravel.org |
 Hagfish (Myxinidae) –°–Ĺ–ł–ľ–ļ–į: Norvan Power Hagfish (Myxinidae) –°–Ĺ–ł–ľ–ļ–į: Norvan Power |
 –ú–ł–Ĺ–ĺ–≥–į (Petromyzontidae) –°–Ĺ–ł–ľ–ļ–į: wikipedia –ú–ł–Ĺ–ĺ–≥–į (Petromyzontidae) –°–Ĺ–ł–ľ–ļ–į: wikipedia |
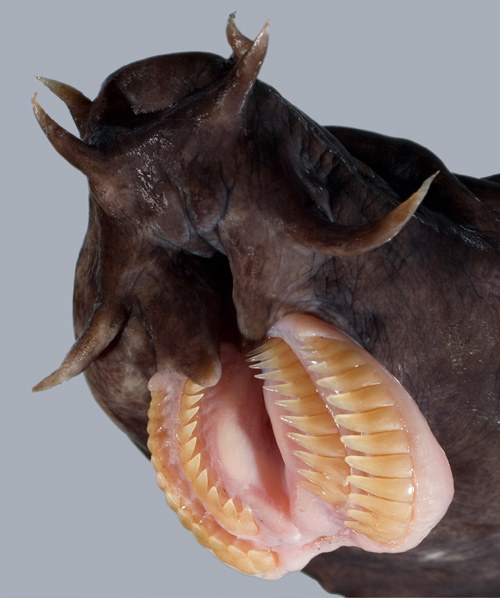 –£—Ā—ā–į—ā–į –Ĺ–į Hagfish (Myxinidae) –°–Ĺ–ł–ľ–ļ–į: nature –£—Ā—ā–į—ā–į –Ĺ–į Hagfish (Myxinidae) –°–Ĺ–ł–ľ–ļ–į: nature |
–ü—Ä–Ķ–∑ –Ņ–ĺ—Ā–Ľ–Ķ–ī–Ĺ–ł—ā–Ķ –ī–Ķ—Ā–Ķ—ā –≥–ĺ–ī–ł–Ĺ–ł –∑–į –ł–∑—Ź—Ā–Ĺ—Ź–≤–į–Ĺ–Ķ—ā–ĺ –Ĺ–į —Ā–Ņ–ĺ—Ä–Ĺ–ł—ā–Ķ –≤—ä–Ņ—Ä–ĺ—Ā–ł –∑–į –Ņ—Ä–ĺ–ł–∑—Ö–ĺ–ī–į –ł —Ä–į–Ĺ–Ĺ–į—ā–į –Ķ–≤–ĺ–Ľ—é—Ü–ł—Ź –Ĺ–į –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—ā–Ķ —Ā–Ķ –≤–ļ–Ľ—é—á–≤–į—ā –į–ļ—ā–ł–≤–Ĺ–ĺ –ł –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ—Ź—Ä–Ĺ–ĺ-–≥–Ķ–Ĺ–Ķ—ā–ł—á–Ĺ–ł –ī–į–Ĺ–Ĺ–ł. –ě—ā–Ĺ–į—á–į–Ľ–ĺ —Ā–į —Ä–į–Ī–ĺ—ā–ł–Ľ–ł —Ā –ĺ—ā–ī–Ķ–Ľ–Ĺ–ł –≥–Ķ–Ĺ–ł, –į —Ā–Ľ–Ķ–ī —ā–ĺ–≤–į –Ĺ–į —Ä–į–∑–Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ–ł–Ķ –Ĺ–į —É—á–Ķ–Ĺ–ł—ā–Ķ —Ā–į –≤–Ķ—á–Ķ –Ņ—ä–Ľ–Ĺ–ł—ā–Ķ —ā–Ķ–ļ—Ā—ā–ĺ–≤–Ķ –Ĺ–į –≥–Ķ–Ĺ–ĺ–ľ–ł—ā–Ķ –Ĺ–į —Ä–į–∑–Ľ–ł—á–Ĺ–ł –≤–ł–ī–ĺ–≤–Ķ –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł, –į —Ā–Ľ–Ķ–ī¬†—ā–ĺ–≤–į –ł —Ü–ł–Ņ–Ķ—Ā—ā–ł. –ē–≤–ĺ–Ľ—é—Ü–ł–ĺ–Ĺ–Ĺ–ł—ā–Ķ –ī—ä—Ä–≤–Ķ—ā–į —Ā—ā–į–≤–į—ā –≤—Ā–Ķ –Ņ–ĺ-—ā–ĺ—á–Ĺ–ł.
–°–Ľ–Ķ–ī –Ņ—Ä–ĺ—á–ł—ā–į–Ĺ–Ķ—ā–ĺ –Ĺ–į –≥–Ķ–Ĺ–ĺ–ľ–į –Ĺ–į –Ľ–į–Ĺ—Ü–Ķ—ā–Ĺ–ł–ļ–į (2008), –Ņ–ĺ—Ā–Ľ–Ķ–ī–Ĺ–ł—Ź –Ľ–ł–Ņ—Ā–≤–į—Č –Ķ–Ľ–Ķ–ľ–Ķ–Ĺ—ā –≤ —Ā–Ľ–ĺ–∂–Ĺ–ł—Ź –Ņ—ä–∑–Ķ–Ľ –Ĺ–į –≤–∑–į–ł–ľ–ĺ–ĺ—ā–Ĺ–ĺ—ą–Ķ–Ĺ–ł—Ź—ā–į –Ĺ–į –≤—ā–ĺ—Ä–ł—á–Ĺ–ĺ—É—Ā—ā–Ĺ–ł—ā–Ķ, –Ķ —É—Ā—ā—Ä–į–Ĺ–ĺ–≤–Ķ–Ĺ–ĺ –ĺ–ļ–ĺ–Ĺ—á–į—ā–Ķ–Ľ–Ĺ–ĺ—ā–ĺ e–≤–ĺ–Ľ—é—Ü–ł–ĺ–Ĺ–Ĺ–ĺ –ī—ä—Ä–≤–ĺ, –ĺ—Ā–Ĺ–ĺ–≤–į–Ĺ–ĺ –Ĺ–į —Ä–į–∑–Ľ–ł—á–ł—Ź—ā–į –≤ –į–ľ–ł–Ĺ–ĺ–ļ–ł—Ā–Ķ–Ľ–ł–Ĺ–Ĺ–ł—ā–Ķ –Ņ–ĺ—Ā–Ľ–Ķ–ī–ĺ–≤–į—ā–Ķ–Ľ–Ĺ–ĺ—Ā—ā–ł –Ĺ–į –Ņ—Ä–ĺ—ā–Ķ–ł–Ĺ–ł—ā–Ķ –≤ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–ł—ā–Ķ:
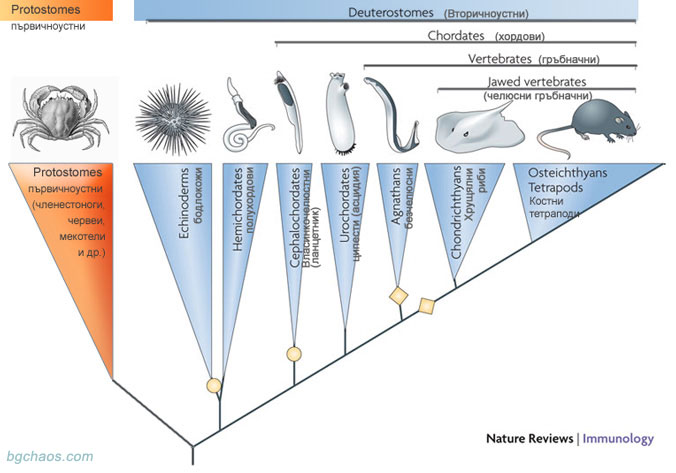 |
| –ě—Ä–ł–≥–ł–Ĺ–į–Ľ–Ĺ–į —Ā—Ö–Ķ–ľ–į: The origins of vertebrate adaptive immunity (–ī–ĺ–Ņ—ä–Ľ–Ĺ–Ķ–Ĺ–į) |
–Ě–į–Ļ-–ī—Ä–Ķ–≤–Ĺ–ł—ā–Ķ —Ā —Ö–ĺ—Ä–ī–į
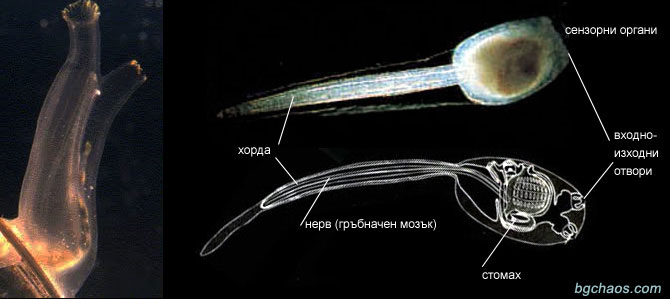
–ö–ĺ—Ź –Ķ —ā–ĺ—á–Ĺ–į—ā–į –ī–į—ā–į –Ĺ–į –Ņ–ĺ—Ź–≤–į—ā–į –Ĺ–į –ĺ–Ī—Č–ł—ā–Ķ –Ņ—Ä–Ķ–ī—Ü–ł –Ĺ–į –ľ–ĺ—Ä—Ā–ļ–ł—ā–Ķ —ā–į—Ä–į–Ľ–Ķ–∂–ł –ł –Ĺ–į –Ĺ–į—Ā, —Ö–ĺ—Ä–į—ā–į, –Ķ —ā—Ä—É–ī–Ĺ–ĺ –ī–į —Ā–Ķ –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–ł, –∑–į—Č–ĺ—ā–ĺ —ā–Ķ —Ā–į –∂–ł–≤–Ķ–Ľ–ł –≤ –ĺ–Ĺ–Ķ–∑–ł –ī—Ä–Ķ–≤–Ĺ–ł –≤—Ä–Ķ–ľ–Ķ–Ĺ–į, –ļ–ĺ–≥–į—ā–ĺ –ĺ—Č–Ķ –Ĺ–Ķ –Ķ –Ī–ł–Ľ –ł–∑–ĺ–Ī—Ä–Ķ—ā–Ķ–Ĺ —Ā–ļ–Ķ–Ľ–Ķ—ā–į. –Ě–į–Ļ-–Ņ—Ä–ł–ľ–ł—ā–ł–≤–Ĺ–ł—ā–Ķ –Ņ—Ä–Ķ–ī—ą–Ķ—Ā—ā–≤–Ķ–Ĺ–ł—Ü–ł –Ĺ–į —Ä–ł–Ī–ł—ā–Ķ, –į –ł –Ĺ–į –≤—Ā–ł—á–ļ–ł –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł —Ā–į –Ĺ–į–ľ–Ķ—Ä–Ķ–Ĺ–ł –≤ –ļ–į–ľ–Ī—Ä–ł–Ļ—Ā–ļ–ł—ā–Ķ —Ā–Ķ–ī–ł–ľ–Ķ–Ĺ—ā–ł. –Ę–Ķ —Ā–į –Ņ—Ä–ĺ—Ā—ā–ł —Ö–ĺ—Ä–ī–ĺ–≤–ł —Ā —Ü–ł–Ľ–ł–Ĺ–ī—Ä–ł—á–Ĺ–ĺ —ā—Ź–Ľ–ĺ, —Ö—Ä–į–Ĺ–ĺ—Ā–ľ–ł–Ľ–į—ā–Ķ–Ľ–Ĺ–į —Ā–ł—Ā—ā–Ķ–ľ–į –ł –ľ–ł–Ĺ–į–≤–į—Č –Ņ—Ä–Ķ–∑ —Ü—Ź–Ľ–ĺ—ā–ĺ —ā—Ź–Ľ–ĺ –Ĺ–Ķ—Ä–≤–Ķ–Ĺ —Ā—ā–≤–ĺ–Ľ, –Ĺ–į—Ä–Ķ—á–Ķ–Ĺ –Ĺ–ĺ—ā–ĺ—Ö–ĺ—Ä–ī–į.
–ü—Ä–Ķ–∑ –Ņ–ĺ—Ā–Ľ–Ķ–ī–Ĺ–ł—ā–Ķ –≥–ĺ–ī–ł–Ĺ–ł –ļ–ł—ā–į–Ļ—Ā–ļ–ł—ā–Ķ –Ņ–į–Ľ–Ķ–ĺ–Ĺ—ā–ĺ–Ľ–ĺ–∑–ł —Ā–į –ĺ—ā–ļ—Ä–ł–Ľ–ł –ł–∑–ļ–ĺ–Ņ–į–Ķ–ľ–ł –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–ł —É—á—É–ī–≤–į—Č–ĺ –ī–ĺ–Ī—Ä–Ķ –∑–į–Ņ–į–∑–Ķ–Ĺ–ł, –ī–ĺ—Ä–ł —Ā –ĺ—ā–Ņ–Ķ—á–į—ā—ä—Ü–ł –Ĺ–į —Ā—ā—Ä—É–ļ—ā—É—Ä–ł—ā–Ķ, –ĺ–Ī—Ä–į–∑—É–≤–į–Ĺ–ł –ĺ—ā –ľ–Ķ–ļ–ł—ā–Ķ —ā—ä–ļ–į–Ĺ–ł. –Ě–į–ľ–Ķ—Ä–Ķ–Ĺ–ł —Ā–į –≤ —Ü–Ķ–Ĺ—ā—Ä–į–Ľ–Ĺ–į—ā–į —á–į—Ā—ā –Ĺ–į –ģ–Ĺ–į–Ĺ (–ģ–∂–Ķ–Ĺ –ö–ł—ā–į–Ļ) –≤ –ī–ĺ–Ľ–Ĺ–ĺ–ļ–į–ľ–Ī—Ä–ł–Ļ—Ā–ļ–ł—ā–Ķ —ą–ł—Ā—ā–ł —Ą–ĺ—Ä–ľ–ł—Ä–į–Ľ–ł –ú–į–ĺ—ā—Ź–Ĺ—ą–į–Ĺ, —á–ł—Ź—ā–ĺ –≤—ä–∑—Ä–į—Ā—ā –Ķ –ĺ–ļ–ĺ–Ľ–ĺ 530 –ľ–ł–Ľ–ł–ĺ–Ĺ–į –≥–ĺ–ī–ł–Ĺ–ł.
Yunnanozoon
–ě—Ä–≥–į–Ĺ–ł–∑–ľ—ä—ā, —á–ł–ł—ā–ĺ –ĺ—Ā—ā–į–Ĺ–ļ–ł –Ī—Ź—Ö–į –ĺ—ā–ļ—Ä–ł—ā–ł –Ņ—Ä–Ķ–∑ 1995 –≥–ĺ–ī–ł–Ĺ–į –≤ –®–Ķ–Ĺ—Ź–Ĺ, –Ķ –Ĺ–į—Ä–Ķ—á–Ķ–Ĺ —Ā –ł–ľ–Ķ—ā–ĺ –Ĺ–į –Ņ—Ä–ĺ–≤–ł–Ĺ—Ü–ł—Ź—ā–į - —é–Ĺ–į–Ĺ–ĺ–∑–ĺ–ĺ–Ĺ, (Yunnanozoon) (Chen J.-Y. et al. // Nature. 1995. V.377. P.720-722). –Ě—Ź–ļ–ĺ–ł –Ķ–ļ—Ā–Ņ–Ķ—Ä—ā–ł –≥–ĺ —Ā–ľ—Ź—ā–į—ā –∑–į –Ĺ–į–Ļ-—Ā—ā–į—Ä–ł—Ź—ā –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–ł—ā–Ķ–Ľ –Ī–Ķ–∑—á–Ķ—Ä–Ķ–Ņ–Ĺ–ł—ā–Ķ —Ö–ĺ—Ä–ī–ĺ–≤–ł, (Acrania), –ī—Ä—É–≥–ł –≥–ĺ –Ņ—Ä–ł—á–ł—Ā–Ľ—Ź–≤–į—ā –ļ—ä–ľ –Ņ–ĺ–Ľ—É—Ö–ĺ—Ä–ī–ĺ–≤–ł—ā–Ķ (Hemichordata).
 |
| Yunnanozoon. –Į—Ā–Ĺ–ĺ —Ā–Ķ –∑–į–Ī–Ķ–Ľ—Ź–∑–≤–į—ā —Ö—Ä–ł–Ľ–Ķ –ł –ļ—É—ā–ł–ļ—É–Ľ—Ź—Ä–Ĺ–ł —Ā–Ķ–≥–ľ–Ķ–Ĺ—ā–ł, —Ä–į–∑–Ľ–ł—á–Ĺ–ł –ĺ—ā –ľ–ł–ĺ–ľ–Ķ—Ä–ł—ā–Ķ (–ľ—É—Ā–ļ—É–Ľ–Ĺ–ł —Ā—ā—Ä—É–ļ—ā—É—Ä–ł) –Ĺ–į —Ö–ĺ—Ä–ī–ĺ–≤–ł—ā–Ķ (–Ņ–ĺ –≥—ä—Ä–Ī–į). –ė–∑–ĺ–Ī—Ä–į–∂–Ķ–Ĺ–ł—Ź –ĺ—ā —Ā–į–Ļ—ā–ĺ–≤–Ķ: bioteaching, geo.tu-freiberg.de |
–° –ľ–Ĺ–ĺ–≥–ĺ –Ņ–ĺ-–≥–ĺ–Ľ—Ź–ľ–į –≤–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ—Ā—ā —Ā–Ķ –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ—Ź –ļ–į—ā–ĺ —Ö–ĺ—Ä–ī–ĺ–≤–į –ī—Ä—É–≥–į –ļ–ł—ā–į–Ļ—Ā–ļ–į –Ĺ–į—Ö–ĺ–ī–ļ–į:
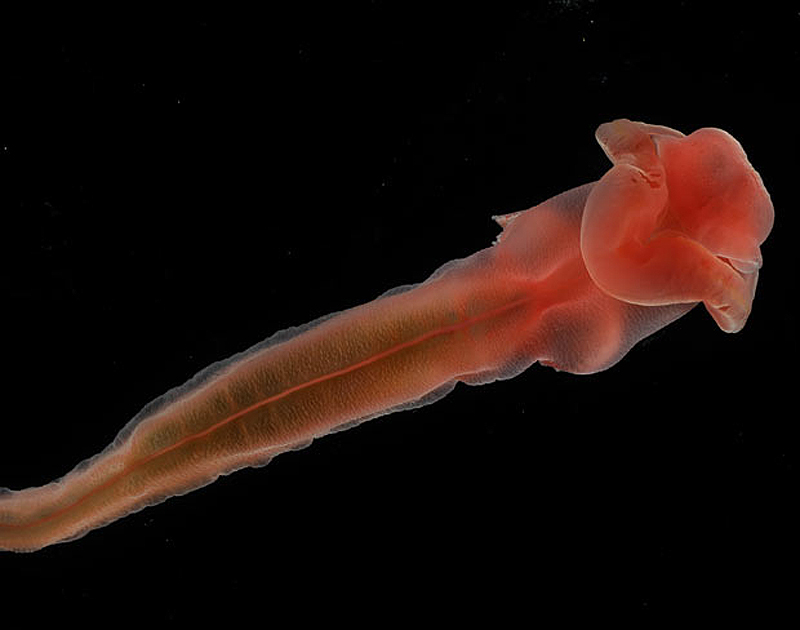
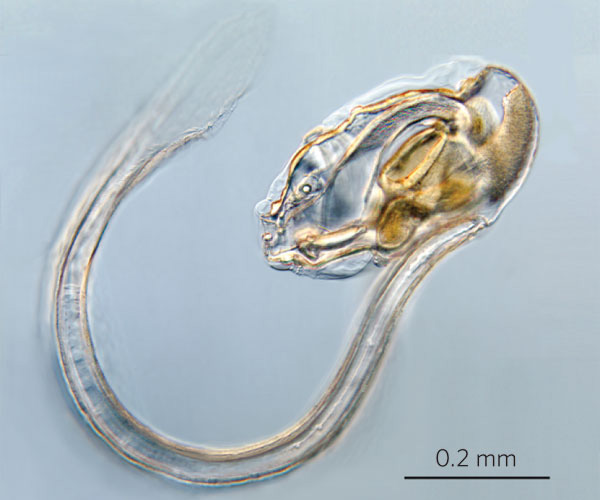
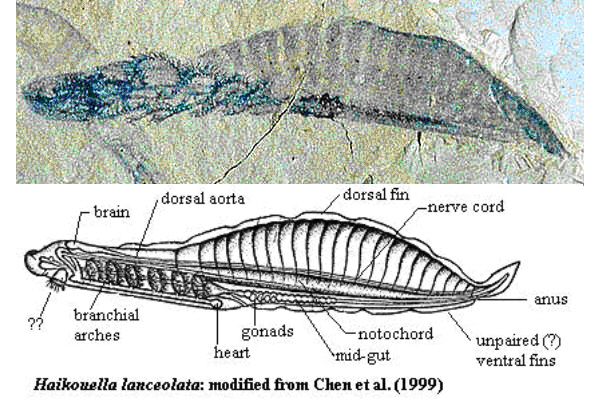
Haikouella
–ü—Ä–Ķ–∑ 1999–≥. –≤ –•–į–Ļ–ļ–ĺ—É, –Ī–Ľ–ł–∑–ĺ –ī–ĺ –ö—É–Ĺ–ľ–ł–Ĺ, –Ī–ł–Ľ–ĺ —Ä–į–∑–ļ–ĺ–Ņ–į–Ĺ–ĺ —Ü—Ź–Ľ–ĺ –≥—Ä–ĺ–Ī–ł—Č–Ķ - 305 –Ķ–ļ–∑–Ķ–ľ–Ņ–Ľ—Ź—Ä–į –Ĺ–į –ī—Ä—É–≥–ĺ —Ö–ĺ—Ä–ī–ĺ–≤–ĺ –∂–ł–≤–ĺ—ā–Ĺ–ĺ, –Ĺ–į—Ä–Ķ—á–Ķ–Ĺ–ĺ –•–į–Ļ–ļ–ĺ—É–Ķ–Ľ–į (Haikouella). –Ę–ĺ–≤–į –ł–∑–ĺ–Ī–ł–Ľ–ł–Ķ –ł –ī–ĺ–Ī—Ä–į—ā–į —Ā—ä—Ö—Ä–į–Ĺ–Ķ–Ĺ–ĺ—Ā—ā –Ņ–ĺ–ľ–ĺ–≥–Ĺ–į–Ľ–ł –Ĺ–į –Ņ–į–Ľ–Ķ–ĺ–Ĺ—ā–ĺ–Ľ–ĺ–∑–ł—ā–Ķ –ī–į –≤—ä–∑—Ā—ā–į–Ĺ–ĺ–≤—Ź—ā —Ā—ā—Ä–ĺ–Ķ–∂–į –Ļ —Ā –ī–ĺ—Ā—ā–į—ā—ä—á–Ĺ–į –Ņ—ä–Ľ–Ĺ–ĺ—ā–į(Chen J.-Y. et al. // Nature. 1999. V.402. P.518-521).
 |
| –ė–∑–ĺ–Ī—Ä–į–∂–Ķ–Ĺ–ł–Ķ –ĺ—ā —Ā–į–Ļ—ā–į pigeonchess |
–•–į–Ļ–ļ–ĺ—É–Ķ–Ľ–į—ā–į –Ī–ł–Ľ–į –ī—ä–Ľ–≥–į –ĺ—ā 20 –ī–ĺ 40 –ľ–ľ. –Ę—Ź–Ľ–ĺ—ā–ĺ –Ļ –Ķ –Ī–ł–Ľ–ĺ –Ľ–į–Ĺ—Ü–Ķ—ā–ĺ–≤–ł–ī–Ĺ–ĺ —Ā –ĺ—ā—á–Ķ—ā–Ľ–ł–≤–ł –≥–Ľ–į–≤–į, —ā–ĺ—Ä—Ā –ł –ĺ–Ņ–į—ą–ļ–į.–ě—ā –ī–≤–Ķ—ā–Ķ —Ā—ā—Ä–į–Ĺ–ł –Ĺ–į –≥–Ľ–į–≤–į—ā–į –ł–ľ–į —á–ł—Ą—ā –∑–į–ĺ–Ī–Ľ–Ķ–Ĺ–ł –ĺ–Ī—Ä–į–∑—É–≤–į–Ĺ–ł—Ź, –ļ–ĺ–ł—ā–ĺ —Ā–Ķ –ł–Ĺ—ā–Ķ—Ä–Ņ—Ä–Ķ—ā–ł—Ä–į—ā –ļ–į—ā–ĺ –ĺ—á–ł.
–Ě–į –ī–ĺ–Ľ–Ĺ–į—ā–į —Ā—ā—Ä–į–Ĺ–į –Ĺ–į –≥–Ľ–į–≤–į—ā–į –Ķ —É—Ā—ā–Ĺ–ł—Ź –ĺ—ā–≤–ĺ—Ä, –∑–į–ĺ–Ī–ł–ļ–ĺ–Ľ–Ķ–Ĺ –ĺ—ā –ľ–į–Ľ–ļ–ł –Ņ–ł–Ņ–į–Ľ–į –ļ–į—ā–ĺ –Ņ–ł–Ņ–į–Ľ–į—ā–į –Ĺ–į —Ā—ä–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ł—Ź, –Ĺ–ĺ –Ņ–ĺ-–Ņ—Ä–ł–ľ–ł—ā–ł–≤–Ķ–Ĺ –Ľ–į–Ĺ—Ü–Ķ—ā–Ĺ–ł–ļ. –°–Ľ–Ķ–ī —É—Ā—ā–į—ā–į –Ķ –ĺ–Ī–Ķ–ľ–ł—Ā—ā–ĺ –≥—ä—Ä–Ľ–ĺ, —á–ł–ł—ā–ĺ —Ā—ā—Ä–į–Ĺ–ł—á–Ĺ–ł—ā–Ķ —Ā—ā–Ķ–Ĺ–ł —Ā–į –Ņ—Ä–ĺ–Ĺ–ł–∑–į–Ĺ–ł –ĺ—ā —Ö—Ä–ł–Ľ–Ĺ–ł –ĺ—ā–≤–ĺ—Ä–ł –ł —Ā–Ķ —É–ļ—Ä–Ķ–Ņ–≤–į —Ā —ą–Ķ—Ā—ā —á–ł—Ą—ā–į —Ö—Ä–ł–Ľ–Ĺ–ł –ī—ä–≥–ł. –í—ä–≤ —Ą–į—Ä–ł–Ĺ–ļ—Ā–į –ł–ľ–į –ľ–Ĺ–ĺ–≥–ĺ –ľ–į–Ľ–ļ–ł (0,1 –ľ–ľ) –ĺ–Ī—Ä–į–∑—É–≤–į–Ĺ–ł—Ź, –ļ–ĺ–ł—ā–ĺ –≤–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ —Ā–į –∑—ä–Ī–ł. –•—Ä–į–Ĺ–ĺ—Ā–ľ–ł–Ľ–į—ā–Ķ–Ľ–Ĺ–į—ā–į —Ā–ł—Ā—ā–Ķ–ľ–į –Ķ –Ņ–ĺ –Ņ—Ä–ĺ–ī—ä–Ľ–∂–Ķ–Ĺ–ł–Ķ –Ĺ–į —Ü—Ź–Ľ–ĺ—ā–ĺ —ā—Ź–Ľ–ĺ –ł –∑–į–≤—ä—Ä—ą–≤–į —Ā –į–Ĺ—É—Ā–į –Ĺ–į –≤–Ķ–Ĺ—ā—Ä–į–Ľ–Ĺ–į—ā–į (–ļ–ĺ—Ä–Ķ–ľ–Ĺ–į—ā–į) —Ā—ā—Ä–į–Ĺ–į –Ĺ–į —ā—Ź–Ľ–ĺ—ā–ĺ, –≤ –ĺ—Ā–Ĺ–ĺ–≤–į—ā–į –Ĺ–į –ĺ–Ņ–į—ą–ļ–į—ā–į.
–Ě–į–ī —Ā—ā–ĺ–ľ–į—Ö–į –Ķ —Ä–į–∑–Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ–į —Ö—Ä—É—Č—Ź–Ľ–Ĺ–į —Ö–ĺ—Ä–ī–į. –Ě–į–ī –Ĺ–Ķ—Ź –Ķ –Ĺ–ĺ—ā–ĺ—Ö–ĺ—Ä–ī–į—ā–į - –Ĺ–į–ī–Ľ—ä–∂–Ķ–Ĺ –Ĺ–Ķ—Ä–≤–Ķ–Ĺ —Ā—ā–≤–ĺ–Ľ, –ļ–ĺ–Ļ—ā–ĺ –≤ –≥–Ľ–į–≤–į—ā–į —Ā–Ķ –Ņ—Ä–Ķ–≤—Ä—ä—Č–į –≤ –Ņ—Ä–ł–ľ–ł—ā–ł–≤–Ķ–Ĺ –ľ–ĺ–∑—ä–ļ, –ļ–ĺ–Ļ—ā–ĺ —Ā–Ķ —Ā—ä—Ā—ā–ĺ–ł –ĺ—ā —ā—Ä–ł –ī—Ź–Ľ–į.
–ď–Ľ–į–≤–Ĺ–ł—ā–Ķ –ļ—Ä—ä–≤–ĺ–Ĺ–ĺ—Ā–Ĺ–ł —Ā—ä–ī–ĺ–≤–Ķ - –ļ–ĺ—Ä–Ķ–ľ–Ĺ–į—ā–į –ł –≥—Ä—ä–Ī–Ĺ–į—ā–į –į–ĺ—Ä—ā–į —Ā–į —Ā–≤—ä—Ä–∑–į–Ĺ–ł —Ā —Ö—Ä–ł–Ľ–Ĺ–ł—ā–Ķ –į—Ä—ā–Ķ—Ä–ł–ł, –į —Ā–į–ľ–į—ā–į –ļ–ĺ—Ä–Ķ–ľ–Ĺ–į –į–ĺ—Ä—ā–į –∑–į–Ņ–ĺ—á–≤–į –ĺ—ā –Ĺ–Ķ—Č–ĺ, –Ņ–ĺ–ī–ĺ–Ī–Ĺ–ĺ –Ĺ–į –ľ–Ķ—Ö—É—Ä, –ļ–ĺ–Ķ—ā–ĺ –Ķ –ĺ—á–Ķ–≤–ł–ī–Ĺ–ĺ —Ā—ä—Ä—Ü–Ķ - —ā—Ä—Ź–Ī–≤–į –ī–į –ĺ—ā–Ī–Ķ–Ľ–Ķ–∂–ł–ľ, —á–Ķ —Ā—ä–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ł—Ź—ā –Ľ–į–Ĺ—Ü–Ķ—ā–Ĺ–ł–ļ –Ĺ—Ź–ľ–į —Ā—ä—Ä—Ü–Ķ. –ď—Ä—ä–Ī–Ĺ–į—ā–į —á–į—Ā—ā –Ĺ–į —ā—Ź–Ľ–ĺ—ā–ĺ –Ĺ–į –•–į–Ļ–ļ–ĺ—É–Ķ–Ľ–į—ā–į, –Ĺ–į–ī —Ö–ĺ—Ä–ī–į—ā–į –Ķ –∑–į–Ķ—ā–į –ĺ—ā –ľ–Ķ—ā–į–ľ–Ķ—Ä–Ĺ–ł –ľ—É—Ā–ļ—É–Ľ–ł, —á–ł–ł—ā–ĺ —Ā–Ķ–≥–ľ–Ķ–Ĺ—ā–ł –ľ–ł–ĺ–ľ–Ķ—Ä–ł —Ā–į —Ä–į–∑–ī–Ķ–Ľ–Ķ–Ĺ–ł –ĺ—ā –∑–ľ–ł–Ķ–≤–ł–ī–Ĺ–ł –Ņ—Ä–Ķ–≥—Ä–į–ī–ł - —Ā–Ķ–Ņ—ā–ł. –ė–Ĺ—ā–Ķ—Ä–Ķ—Ā–Ĺ–ĺ –Ķ, —á–Ķ –ľ—É—Ā–ļ—É–Ľ–ł—ā–Ķ –Ĺ–į —Ā—ä–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ł—Ź –Ľ–į–Ĺ—Ü–Ķ—ā–Ĺ–ł–ļ —Ā–į —Ā—ä—Č–ĺ –ľ–Ķ—ā–į–ľ–Ķ—Ä–Ĺ–ł, —ā.–Ķ. –ł–ľ–į —Ā–Ķ–≥–ľ–Ķ–Ĺ—ā–ł—Ä–į–Ĺ–į.
–í—Ā–Ķ –Ņ–į–ļ –Ĺ—Ź–ļ–ĺ–ł –Ņ–į–Ľ–Ķ–ĺ–Ĺ—ā–ĺ–Ľ–ĺ–∑–ł –ĺ—Ā–Ņ–ĺ—Ä–≤–į—ā —ā—ä–Ľ–ļ—É–≤–į–Ĺ–Ķ—ā–ĺ –Ĺ–į –Ĺ—Ź–ļ–ĺ–ł –ī–Ķ—ā–į–Ļ–Ľ–ł –Ĺ–į –ĺ—ā–Ņ–Ķ—á–į—ā—ä—Ü–ł—ā–Ķ –ł –Ņ—Ä–ł—á–ł—Ā–Ľ—Ź–≤–į—ā –•–į–Ļ–ĺ—É–Ķ–Ľ–į—ā–į –ļ—ä–ľ –Ņ–ĺ–Ľ—É—Ö–ĺ—Ä–ī–ĺ–≤–ł—ā–Ķ.
Haikouichthys ercaicunensis
–ě—ā —Ä–į–Ĺ–Ĺ–ł—Ź –ļ–į–ľ–Ī—Ä–ł–Ļ –Ķ –Ķ–ī–Ĺ–ĺ –ī—Ä—É–≥–ĺ –ł–∑–ļ–ĺ–Ņ–į–Ķ–ľ–ĺ –≤–ĺ–ī–Ĺ–ĺ –∂–ł–≤–ĺ—ā–Ĺ–ĺ, –ļ–ĺ–Ķ—ā–ĺ –Ī–Ķ–∑—Ā—ä–ľ–Ĺ–Ķ–Ĺ–ĺ –Ķ –ĺ—ā –Ĺ–į–Ļ-–Ņ—Ä–ł–ľ–ł—ā–ł–≤–Ĺ–ł—ā–Ķ —Ö–ĺ—Ä–ī–ĺ–≤–ł –Ī–Ķ–∑—á–Ķ–Ľ—é—Ā—ā–Ĺ–ł - Haikouichthys ercaicunensis (Shu et al., 2003. Head and backbone of the Early Cambrian vertebrate Haikouichthys).
 –°–Ĺ–ł–ľ–ļ–į: vertebresfossiles.free.fr –°–Ĺ–ł–ľ–ļ–į: vertebresfossiles.free.fr |
–ė–∑–ļ–ĺ–Ņ–į–Ķ–ľ–ł—ā–Ķ –ĺ—Ā—ā–į–Ĺ–ļ–ł –Ĺ–į –•–į–Ļ–ļ—É–ł—Ö—ā–ł—Ā–į –Ī–ł–Ľ–ł –ĺ—ā–ļ—Ä–ł—ā–ł –Ņ—Ä–Ķ–∑ 1999 –≥. –Ī–Ľ–ł–∑–ĺ –ī–ĺ –≥—Ä–į–ī –Ĺ–į –•–į–Ļ–ļ–ĺ—É (–ö–ł—ā–į–Ļ). –ě—ā—ā–ĺ–≥–į–≤–į, —É—á–Ķ–Ĺ–ł—ā–Ķ —Ā–į –ĺ—ā–ļ—Ä–ł–Ľ–ł –Ĺ—Ź–ļ–ĺ–Ľ–ļ–ĺ –ī–Ķ—Ā–Ķ—ā–ļ–ł –ī–ĺ–Ī—Ä–Ķ –∑–į–Ņ–į–∑–Ķ–Ĺ–ł –≤–ļ–į–ľ–Ķ–Ĺ–Ķ–Ľ–ĺ—Ā—ā–ł –Ĺ–į –∂–ł–≤–ĺ—ā–Ĺ–ĺ—ā–ĺ, –ļ–ĺ–Ķ—ā–ĺ –Ņ–ĺ–∑–≤–ĺ–Ľ—Ź–≤–į –ī–Ķ—ā–į–Ļ–Ľ–Ĺ–ĺ –ī–į —Ā–Ķ –ł–∑—É—á–ł —Ā—ā—Ä—É–ļ—ā—É—Ä–į—ā–į –ī–ĺ—Ä–ł –ł –Ĺ–į –ĺ—á–ł—ā–Ķ –ł –≤—ä—ā—Ä–Ķ—ą–Ĺ–ł—ā–Ķ –ľ—É –ĺ—Ä–≥–į–Ĺ–ł.
 –†–Ķ–ļ–ĺ–Ĺ—Ā—ā—Ä—É–ļ—Ü–ł—Ź: Shudong.com –†–Ķ–ļ–ĺ–Ĺ—Ā—ā—Ä—É–ļ—Ü–ł—Ź: Shudong.com |
–ü—Ä–ł–ľ–ł—ā–ł–≤–Ĺ–ł—Ź—ā —Ö—Ä—É—Č—Ź–Ľ–Ķ–Ĺ –≥—Ä—ä–Ī–Ĺ–į–ļ –Ĺ–į Haikouichthys –ľ—É –Ņ–ĺ–∑–≤–ĺ–Ľ—Ź–≤–į –ī–į –Ņ–Ľ—É–≤–į –Ņ–ĺ –ī–ĺ—Ā—ā–į –Ņ–ĺ-—Ä–į–∑–Ľ–ł—á–Ķ–Ĺ –Ĺ–į—á–ł–Ĺ –ĺ—ā —á–Ľ–Ķ–Ĺ–Ķ—Ā—ā–ĺ–Ĺ–ĺ–≥–ł—ā–Ķ. –ú–į–Ľ–ļ–ĺ—ā–ĺ –ľ—É —ā—Ź–Ľ–ĺ (2,5 —Ā–ľ) –Ķ –Ľ–ł—ą–Ķ–Ĺ–ĺ –ĺ—ā –Ņ–Ľ–į–≤–Ĺ–ł—Ü–ł, –Ĺ–ĺ –Ķ –ł–ľ–į–Ľ–ĺ —Ö—Ä–ł–Ľ–Ĺ–ł –ī—ä–≥–ł, –Ĺ–ĺ—Ä–ľ–į–Ľ–Ĺ–ł –ĺ—á–ł –ł –ľ–ĺ–∑—ä–ļ –ļ–į—ā–ĺ –Ĺ—Ź–ļ–ĺ–ł –ĺ—ā –ī–Ķ—ā–į–Ļ–Ľ–ł—ā–Ķ –Ĺ–į –Ņ—Ä–Ķ–ī–Ĺ–ł—Ź –ļ—Ä–į–Ļ –Ĺ–į —ā—Ź–Ľ–ĺ—ā–ĺ –ľ—É –Ĺ–į–Ņ–ĺ–ľ–Ĺ—Ź—ā –≥–Ľ–į–≤–į—ā–į –Ĺ–į Pikaia (–∑–į –ļ–ĺ—Ź—ā–ĺ —Č–Ķ –≥–ĺ–≤–ĺ—Ä–ł–ľ –ľ–į–Ľ–ļ–ĺ –Ņ–ĺ-–Ĺ–į—ā–į—ā—ä–ļ).
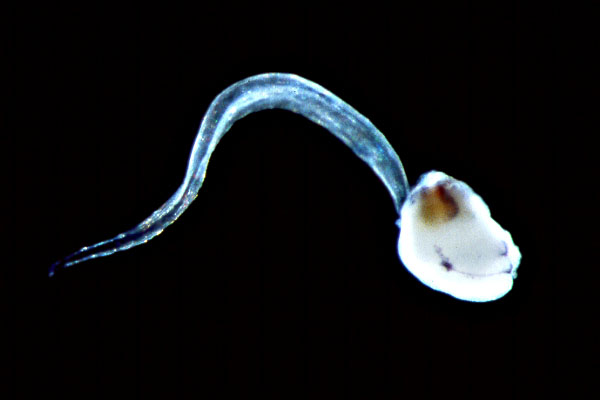
Myllokunmingia
Myllokunmingia –Ķ —Ö–ĺ—Ä–ī–ĺ–≤–ĺ –ĺ—ā –Ē–ĺ–Ľ–Ĺ–ł—Ź –ö–į–ľ–Ī—Ä–ł–Ļ, –Ĺ–į–ľ–Ķ—Ä–Ķ–Ĺ–ĺ –≤ —ą–ł—Ā—ā–ł—ā–Ķ –ú–į–ĺ—ā—Ź–Ĺ—ą–į–Ĺ (–ö–ł—ā–į–Ļ). –°–ľ—Ź—ā–į —Ā–Ķ, —á–Ķ –Ķ –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ĺ, –Ĺ–ĺ –∑–į—Ā–Ķ–≥–į —ā–ĺ–≤–į –Ĺ–Ķ –Ķ –ī–ĺ–ļ–į–∑–į–Ĺ–ĺ —É–Ī–Ķ–ī–ł—ā–Ķ–Ľ–Ĺ–ĺ.
 –°–Ĺ–ł–ľ–ļ–į: vertebresfossiles.free.fr –°–Ĺ–ł–ľ–ļ–į: vertebresfossiles.free.fr |
Myllokunmingia –Ķ —Ā –ī—ä–Ľ–∂–ł–Ĺ–į 28 –ľ–ľ –ł 6 –ľ–ľ. –Ę—Ź –Ķ —Ā—Ä–Ķ–ī –Ĺ–į–Ļ-—Ā—ā–į—Ä–ł—ā–Ķ –Ĺ–į –≤—ä–∑–ľ–ĺ–∂–Ĺ–ł —á–Ķ—Ä–Ķ–Ņ–Ĺ–ł, –Ĺ–į–ľ–Ķ—Ä–Ķ–Ĺ–ł –≤ –Ē–ĺ–Ľ–Ĺ–ł—Ź –ö–į–ľ–Ī—Ä–ł–Ļ –Ņ—Ä–Ķ–ī–ł 524 –ľ–ł–Ľ–ł–ĺ–Ĺ–į –≥–ĺ–ī–ł–Ĺ–ł.¬†
 |
–Ě–į –ĺ—ā–Ņ–Ķ—á–į—ā—ä–ļ—ä—ā –Ľ–ł—á–ł —á–Ķ—Ä–Ķ–Ņ –ł —Ā–ļ–Ķ–Ľ–Ķ—ā, –Ĺ–į–Ņ—Ä–į–≤–Ķ–Ĺ–ł –ĺ—ā —Ö—Ä—É—Č—Ź–Ľ. –Ė–ł–≤–ĺ—ā–Ĺ–ĺ—ā–ĺ –ł–ľ–į –ĺ–Ī–ĺ—Ā–ĺ–Ī–Ķ–Ĺ–į –≥–Ľ–į–≤–į –ł —ā—Ź–Ľ–ĺ. –ď–Ľ–į–≤–į—ā–į –ł–ľ–į –Ņ–Ķ—ā –ł–Ľ–ł —ą–Ķ—Ā—ā —Ö—Ä–ł–Ľ–Ĺ–ł —ā–ĺ—Ä–Ī–ł—á–ļ–ł. –ü–ĺ —ā—Ź–Ľ–ĺ—ā–ĺ —Ā–ł –ł–ľ–į –ĺ—Ä–ł–Ķ–Ĺ—ā–ł—Ä–į–Ĺ–ł –Ĺ–į–∑–į–ī, V-–ĺ–Ī—Ä–į–∑–Ĺ–ł 25 —Ā–Ķ–≥–ľ–Ķ–Ĺ—ā–ł - –ľ–ł–ĺ–ľ–Ķ—Ä–ł. –Ě–į–Ľ–ł—Ü–Ķ –Ķ –Ĺ–ĺ—ā–ĺ—Ö–ĺ—Ä–ī–į, —É—Ā—ā–Ĺ–į –ļ—É—Ö–ł–Ĺ–į –ł —Ö—Ä–į–Ĺ–ĺ—Ā–ľ–ł–Ľ–į—ā–Ķ–Ľ–Ķ–Ĺ —ā—Ä–į–ļ—ā, —Ä–į–∑–Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ –Ņ–ĺ –Ņ—Ä–ĺ–ī—ä–Ľ–∂–Ķ–Ĺ–ł–Ķ –Ĺ–į —Ü—Ź–Ľ–ĺ—ā–ĺ —ā—Ź–Ľ–ĺ –Ĺ–į –∂–ł–≤–ĺ—ā–Ĺ–ĺ—ā–ĺ. –Ě—Ź–ľ–į —Ä–į–ī–ł–į–Ľ–Ĺ–ł –Ņ–Ķ—Ä–ļ–ł.
–ú–į–Ľ–ļ–ĺ –Ņ–ĺ-–ļ—ä—Ā–Ĺ–ĺ, —Ā–Ķ –ĺ—ā–ī–Ķ–Ľ—Ź –≥—Ä—É–Ņ–į—ā–į –Ĺ–į —Ü–Ķ—Ą–į–Ľ–ĺ—Ö–ĺ—Ä–ī–ĺ–≤–ł—ā–Ķ, –ĺ—ā–Ľ–ł—á–į–≤–į—Č–ł —Ā–Ķ —Ā –ĺ–Ņ–į—ą–ļ–į –ł –≥—Ä—É–Ņ–į –Ĺ–į –Ĺ–Ķ—Ä–≤–ł –≤ –Ņ—Ä–Ķ–ī–Ĺ–į—ā–į —á–į—Ā—ā –Ĺ–į —ā—Ź–Ľ–ĺ—ā–ĺ. –°—Ä–Ķ–ī —ā—Ź—Ö –Ķ:
Pikaia gracilens
–í –Ņ–ĺ-–ļ—ä—Ā–Ĺ–ł—ā–Ķ –ĺ—ā –ļ–ł—ā–į–Ļ—Ā–ļ–ł—ā–Ķ, –ļ–į–ľ–Ī—Ä–ł–Ļ—Ā–ļ–ł —Ā–Ķ–ī–ł–ľ–Ķ–Ĺ—ā–ł –Ĺ–į –Ď—ä—Ä–ī–∂–Ķ—Ā –®–Ķ–Ļ–Ľ (Burgess Shal–Ķ) —Ā—ä—Č–ĺ —Ā–į –Ĺ–į–ľ–Ķ—Ä–Ķ–Ĺ–ł –ĺ—ā–Ņ–Ķ—á–į—ā—ä—Ü–ł –ĺ—ā —Ä–į–∑–Ĺ–ĺ–ĺ–Ī—Ä–į–∑–Ĺ–ł –ľ–Ķ–ļ–ĺ—ā–Ķ–Ľ–ł (–Ī–Ķ–∑ —Ā–ļ–Ķ–Ľ–Ķ—ā –ł–Ľ–ł –Ĺ—Ź–ļ–į–ļ–≤–į —ā–≤—ä—Ä–ī–į –ĺ–Ī–≤–ł–≤–ļ–į) –∂–ł–≤–ĺ—ā–Ĺ–ł, –Ņ—Ä–ł–Ľ–ł—á–į—Č–ł –Ĺ–į –Ņ—Ä–ł–ľ–ł—ā–ł–≤–Ĺ–ł —Ö–ĺ—Ä–ī–ĺ–≤–ł.
–°—ä–Ī—Ä–į–Ĺ –Ķ –ī–ĺ—Ā—ā–į—ā—ä—á–Ķ–Ĺ –ľ–į—ā–Ķ—Ä–ł–į–Ľ –∑–į –ī–į –ľ–ĺ–≥–į—ā –Ņ–į–Ľ–Ķ–ĺ–Ĺ—ā–ĺ–Ľ–ĺ–∑–ł—ā–Ķ –ī–į —Ä–Ķ–ļ–ĺ–Ĺ—Ā—ā—Ä—É–ł—Ä–į—ā –Ņ–ĺ–ī—Ä–ĺ–Ī–Ĺ–ĺ –į–Ĺ–į—ā–ĺ–ľ–ł—Ź—ā–į –Ĺ–į –Ķ–ī–Ĺ–ĺ —Ā—ä—Č–Ķ—Ā—ā–≤–ĺ - Pikaia gracilens.
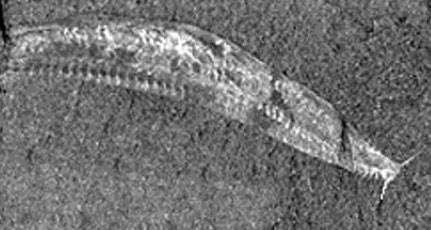 –°–Ĺ–ł–ľ–ļ–į (—Ą—Ä–į–≥–ľ–Ķ–Ĺ—ā): Smithsonian Institution –°–Ĺ–ł–ľ–ļ–į (—Ą—Ä–į–≥–ľ–Ķ–Ĺ—ā): Smithsonian Institution |
Pikaia gracilens —Ā–Ķ –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ—Ź –ļ–į—ā–ĺ –Ĺ–į–Ļ-"–Ī–į–∑–į–Ľ–Ĺ–ĺ", (–Ĺ–į–Ļ-–Ņ—Ä–ł–ľ–ł—ā–ł–≤–Ĺ–ĺ) –ĺ—ā –Ņ–ĺ–∑–Ĺ–į—ā–ł—ā–Ķ –ī–ĺ –ľ–ĺ–ľ–Ķ–Ĺ—ā–į —Ö–ĺ—Ä–ī–ĺ–≤–ł. |
Pikaia —Ā–Ķ —Ä–į–∑–Ľ–ł—á–į–≤–į –ĺ—ā –≤—Ā–ł—á–ļ–ł –ł–∑–≤–Ķ—Ā—ā–Ĺ–ł –Ĺ–ł—Ā—ą–ł —Ö–ĺ—Ä–ī–ĺ–≤–ł –Ņ–ĺ –Ĺ—Ź–ļ–ĺ–ł —É–Ĺ–ł–ļ–į–Ľ–Ĺ–ł –Ņ—Ä–ł–∑–Ĺ–į—Ü–ł. –Ę—Ź –Ķ –ł–ľ–į–Ľ–į –ľ–į–Ľ–ļ–į –Ņ–Ľ–ĺ—Ā–ļ–į –≥–Ľ–į–≤–į —Ā —á–ł—Ą—ā –ī—ä–Ľ–≥–ł –Ņ–ł–Ņ–į–Ľ–į –ł –ī–Ķ–≤–Ķ—ā –ī–≤–ĺ–Ļ–ļ–ł —Ä–į–∑–ļ–Ľ–ĺ–Ĺ–Ķ–Ĺ–ł –ł–∑–ī–į—ā—ä—Ü–ł –Ĺ–į –Ņ—Ä–Ķ–ī–Ĺ–ł—Ź –ļ—Ä–į–Ļ –Ĺ–į —ā—Ź–Ľ–ĺ—ā–ĺ (–≤–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ –≤—ä–Ĺ—ą–Ĺ–ł —Ö—Ä–ł–Ľ–Ķ). –í –ĺ—Ā–Ĺ–ĺ–≤–į—ā–į –Ĺ–į —ā–Ķ–∑–ł –ł–∑–ī–į—ā—ä—Ü–ł –ł–ľ–į –ľ–į–Ľ–ļ–ł –ī—É–Ņ–ļ–ł, —Ā–≤—ä—Ä–∑–≤–į—Č–ł —É—Ā—ā–Ĺ–į—ā–į –ļ—É—Ö–ł–Ĺ–į —Ā –ĺ–ļ–ĺ–Ľ–Ĺ–į—ā–į —Ā—Ä–Ķ–ī–į. –ė–ľ–į–Ľ–į –Ķ –≥–ĺ—Ä–Ĺ–į –ł –ī–ĺ–Ľ–Ĺ–į —ā–Ķ—Ā–Ĺ–ł –Ņ–Ķ—Ä–ļ–ł. –Ē—ä–Ľ–≥–ĺ—ā–ĺ, —Ā–Ņ–Ľ–Ķ—Ā–ļ–į–Ĺ–ĺ —Ā—ā—Ä–į–Ĺ–ł—á–Ĺ–ĺ —ā—Ź–Ľ–ĺ –Ĺ–į Pikaia –ł–ľ–į —Ö–į—Ä–į–ļ—ā–Ķ—Ä–Ĺ–ł—ā–Ķ —Ā–į–ľ–ĺ –∑–į —Ö–ĺ—Ä–ī–ĺ–≤–ł—ā–Ķ –ł–∑–≤–ł—ā–ł –ľ—É—Ā–ļ—É–Ľ–Ĺ–ł —Ā–Ķ–≥–ľ–Ķ–Ĺ—ā–ł - –ľ–ł–ĺ–ľ–Ķ—Ä–ł –ł —Ö–ĺ—Ä–ī–į.
 –†–Ķ–ļ–ĺ–Ĺ—Ā—ā—Ä—É–ļ—Ü–ł—Ź: Phlesch Bubble (–Ņ—Ä–Ķ—Ä–į–Ī–ĺ—ā–Ķ–Ĺ–į) –†–Ķ–ļ–ĺ–Ĺ—Ā—ā—Ä—É–ļ—Ü–ł—Ź: Phlesch Bubble (–Ņ—Ä–Ķ—Ä–į–Ī–ĺ—ā–Ķ–Ĺ–į) |
–ü–ĺ–Ľ—É—á–Ķ–Ĺ–ł—ā–Ķ –ĺ—ā –Ņ–į–Ľ–Ķ–ĺ–Ĺ—ā–ĺ–Ľ–ĺ–∑–ł—ā–Ķ –ī–į–Ĺ–Ĺ–ł —Ā–į –≤ —Ā—ä–ĺ—ā–≤–Ķ—ā—Ā—ā–≤–ł–Ķ —Ā —Ö–ł–Ņ–ĺ—ā–Ķ–∑–į—ā–į –∑–į –Ņ—Ä–ĺ–ł–∑—Ö–ĺ–ī–į –Ĺ–į —Ö–ĺ—Ä–ī–ĺ–≤–ł—ā–Ķ –ĺ—ā –ī—Ä–Ķ–≤–Ĺ–ł—ā–Ķ –ľ–ĺ—Ä—Ā–ļ–ł —á–Ķ—Ä–≤–Ķ–ł, –ļ–ĺ–ł—ā–ĺ —Ā–į –Ī–Ľ–ł–∑–ĺ –ī–ĺ –Ĺ–ł–≤–ĺ—ā–ĺ –Ĺ–į –ĺ—Ä–≥–į–Ĺ–ł–∑–į—Ü–ł—Ź –Ĺ–į —Ā—ä–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ł—ā–Ķ –Ņ–ĺ–Ľ—É—Ö–ĺ—Ä–ī–ĺ–≤–ł (Enteropneusta). –ú–ĺ–∂–Ķ –Ī–ł –≤ –Ĺ–į–Ļ-—Ä–į–Ĺ–Ĺ–ł—ā–Ķ –Ķ—ā–į–Ņ–ł –Ĺ–į –Ķ–≤–ĺ–Ľ—é—Ü–ł—Ź—ā–į –Ĺ–į —Ö–ĺ—Ä–ī–ĺ–≤–ł—ā–Ķ, —É —ā–Ķ—Ö–Ĺ–ł—ā–Ķ –Ņ—Ä–Ķ–ī—Ü–ł —Ā–į —Ā–Ķ —Ä–į–∑–≤–ł–Ľ–ł –Ĺ–ĺ–≤–ł –Ņ—Ä–ł—Ā–Ņ–ĺ—Ā–ĺ–Ī–Ľ–Ķ–Ĺ–ł—Ź (–Ĺ–ĺ—ā–ĺ—Ö–ĺ—Ä–ī–į, —Ā–Ķ–≥–ľ–Ķ–Ĺ—ā–ł—Ä–į–Ĺ–Ķ –Ĺ–į –ľ—É—Ā–ļ—É–Ľ–ł—ā–Ķ –Ĺ–į —ā—Ź–Ľ–ĺ—ā–ĺ, –Ņ–ĺ-—Ā–Ľ–ĺ–∂–Ĺ–ł —Ā–Ķ—ā–ł–≤–Ĺ–ł –ĺ—Ä–≥–į–Ĺ–ł –ł —Ü–Ķ–Ĺ—ā—Ä–į–Ľ–Ĺ–į—ā–į –Ĺ–Ķ—Ä–≤–Ĺ–į —Ā–ł—Ā—ā–Ķ–ľ–į).
–Ę–Ķ–∑–ł –Ĺ–ĺ–≤–ĺ–≤—ä–≤–Ķ–ī–Ķ–Ĺ–ł—Ź —Ā–į –ł–ľ –ī–į–Ľ–ł –≤—ä–∑–ľ–ĺ–∂–Ĺ–ĺ—Ā—ā –ī–į –Ņ—Ä–Ķ–ľ–ł–Ĺ–į—ā –ĺ—ā –Ņ—Ä–ł–ī—ä–Ĺ–Ķ–Ĺ –Ĺ–į—á–ł–Ĺ –Ĺ–į –∂–ł–≤–ĺ—ā –ł —á–Ķ—Ä–≤–Ķ–ĺ–Ī—Ä–į–∑–Ķ–Ĺ –Ĺ–į—á–ł–Ĺ –Ĺ–į –Ņ—Ä–ł–ī–≤–ł–∂–≤–į–Ĺ–Ķ –ļ—ä–ľ —Ā–≤–ĺ–Ī–ĺ–ī–Ĺ–ĺ –į–ļ—ā–ł–≤–Ĺ–ĺ –Ņ–Ľ—É–≤–į–Ĺ–Ķ. –ó–į–Ņ–į–∑–ł–Ľ–ł —Ā–į —Ā—ā–į—Ä–ł—Ź—ā –Ĺ–į—á–ł–Ĺ –Ĺ–į —Ö—Ä–į–Ĺ–Ķ–Ĺ–Ķ –Ĺ–į –ľ–ĺ—Ä—Ā–ļ–ł—ā–Ķ —á–Ķ—Ä–≤–Ķ–ł - —Ą–ł–Ľ—ā—Ä–ł—Ä–į–Ĺ–Ķ –Ĺ–į –ľ–ł–ļ—Ä–ĺ–ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–ł –ł —á–į—Ā—ā–ł—Ü–ł —É—ā–į–Ļ–ļ–į —Ā —É—Ā—ā–į—ā–į. –ú–ĺ–∂–Ķ –Ī–ł –≤ –ĺ–Ĺ–Ķ–∑–ł –≤—Ä–Ķ–ľ–Ķ–Ĺ–į —Ö–ĺ—Ä–ī–ĺ–≤–ł—ā–Ķ —Ā–į —Ā–Ķ —Ä–į–∑–ī–Ķ–Ľ–ł–Ľ–ł –Ĺ–į —ā—Ä–ł –≥—Ä—É–Ņ–ł, –ļ–ĺ–ł—ā–ĺ —Ā–Ķ —Ä–į–∑–ī–Ķ–Ľ—Ź—ā –≤ —Ö–ĺ–ī–į –Ĺ–į –Ķ–≤–ĺ–Ľ—é—Ü–ł—Ź—ā–į –Ĺ–į —ā—Ä–ł—ā–Ķ —Ā—ä–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ł –Ņ–ĺ–ī—ā–ł–Ņ–į: —Ü–ł–Ņ–Ķ—Ā—ā–ł, –Ī–Ķ–∑—á–Ķ—Ä–Ķ–Ņ–Ĺ–ł –ł –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł –∂–ł–≤–ĺ—ā–Ĺ–ł.
–°—ä–ī–Ķ–Ļ–ļ–ł –Ņ–ĺ –ĺ–Ņ–ł—Ā–į–Ĺ–ł—ā–Ķ –Ĺ–į—Ö–ĺ–ī–ļ–ł, —Ö–ĺ—Ä–ī–ĺ–≤–ł—ā–Ķ —Ā–Ķ –ĺ—Ą–ĺ—Ä–ľ—Ź—ā –≤ –Ĺ–į—á–į–Ľ–ĺ—ā–ĺ –Ĺ–į –Ņ–Ķ—Ä–ł–ĺ–ī–į –ļ–į–ľ–Ī—Ä–ł–Ļ - –Ņ–ĺ –≤—Ä–Ķ–ľ–Ķ –Ĺ–į "–ö–į–ľ–Ī—Ä–ł–Ļ—Ā–ļ–ł—Ź –≤–∑—Ä–ł–≤", –Ĺ–ĺ –ļ–į–ļ—ā–ĺ –Ņ–ĺ–ļ–į–∑–≤–į—ā –Ņ–ĺ—Ā–Ľ–Ķ–ī–Ĺ–ł—ā–Ķ –ł–∑—Ā–Ľ–Ķ–ī–≤–į–Ĺ–ł—Ź –Ĺ–į –Ē–Ě–ö, –Ņ—Ä–į–ī–Ķ–ī–ł—ā–Ķ –Ĺ–į –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—ā–Ķ –ł–∑–≥–Ľ–Ķ–∂–ī–į –ī–į —Ā–į —Ā–Ķ —Ä–į–∑–ī–Ķ–Ľ–ł–Ľ–ł –ĺ—ā –Ī–Ķ–∑–≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł—ā–Ķ –ĺ—Č–Ķ –≤ –Ē–ĺ–ļ–į–ľ–Ī—Ä–ł–Ļ, –Ņ—Ä–Ķ–ī–ł –ĺ–ļ–ĺ–Ľ–ĺ 600 –ľ–ł–Ľ–ł–ĺ–Ĺ–į –≥–ĺ–ī–ł–Ĺ–ł.
–§–į–ļ—ā—ä—ā, —á–Ķ –Ī–į–∑–į–Ľ–Ĺ–ł –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł –ļ–į—ā–ĺ –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ķ–Ĺ–ł—ā–Ķ —Ā–Ķ –Ņ–ĺ—Ź–≤—Ź–≤–į—ā –≤—ä–≤ —Ą–ĺ—Ā–ł–Ľ–Ĺ–ł—ā–Ķ –ī–į–Ĺ–Ĺ–ł, –ľ–Ĺ–ĺ–≥–ĺ –Ņ—Ä–Ķ–ī–ł –ł—Ā—ā–ł–Ĺ—Ā–ļ–ł—ā–Ķ –≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł –∂–ł–≤–ĺ—ā–Ĺ–ł –ł —Ā–į –ľ–ĺ—Ä—Ą–ĺ–Ľ–ĺ–≥–ł—á–Ĺ–ĺ –Ņ–ĺ–ī–ĺ–Ī–Ĺ–ł –Ķ–ī–Ĺ–ĺ–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ –ł –Ĺ–į —Ö–ĺ—Ä–ī–ĺ–≤–ł –ł –Ĺ–į –Ī–Ķ–∑–≥—Ä—ä–Ī–Ĺ–į—á–Ĺ–ł –Ķ —ā–ĺ—á–Ĺ–ĺ —ā–ĺ–≤–į, –ļ–ĺ–Ķ—ā–ĺ —á–ĺ–≤–Ķ–ļ –Ī–ł –ĺ—á–į–ļ–≤–į–Ľ, —Ā—ä–≥–Ľ–į—Ā–Ĺ–ĺ –Ķ–≤–ĺ–Ľ—é—Ü–ł–ĺ–Ĺ–Ĺ–į—ā–į —ā–Ķ–ĺ—Ä–ł—Ź.
–ü—Ä–ł–Ľ–ĺ–∂–Ķ–Ĺ–ł–Ķ 1: –§–ł–Ľ–ľ –∑–į –ē–≤–ĺ–Ľ—é—Ü–ł—Ź—ā–į –Ĺ–į –•–ĺ—Ä–ī–ĺ–≤–ł—ā–Ķ - —á–į—Ā—ā 1
–ü—Ä–ł–Ľ–ĺ–∂–Ķ–Ĺ–ł–Ķ 2: –Ę–į–Ī–Ľ–ł—Ü–į –Ĺ–į –≥–Ķ–ĺ–Ľ–ĺ–∂–ļ–ł—ā–Ķ –Ņ–Ķ—Ä–ł–ĺ–ī–ł
(o—ā –í–ł–ļ–ł–Ņ–Ķ–ī–ł—Ź)
| –ē–ĺ–Ĺ | –ē—Ä–į –ü—Ä–ĺ–ī—ä–Ľ–∂–ł—ā–Ķ–Ľ–Ĺ–ĺ—Ā—ā | –ü–Ķ—Ä–ł–ĺ–ī | –Ě–į—á–į–Ľ–ĺ –≤ –ľ–Ľ–Ĺ. –≥. |
|---|---|---|---|
| –§–į–Ĺ–Ķ—Ä–ĺ–∑–ĺ–Ļ | |||
| –Ě–Ķ–ĺ–∑–ĺ–Ļ 65,5 –ľ–Ľ–Ĺ. –≥. | –ö–≤–į—ā–Ķ—Ä–Ĺ–Ķ—Ä | 2,0 | |
| –Ě–Ķ–ĺ–≥–Ķ–Ĺ | 23,03 | ||
| –ü–į–Ľ–Ķ–ĺ–≥–Ķ–Ĺ | 65,5 | ||
| –ú–Ķ–∑–ĺ–∑–ĺ–Ļ 185,5 –ľ–Ľ–Ĺ. –≥. | –ö—Ä–Ķ–ī–į | 145,5 | |
| –ģ—Ä–į | 199,6 | ||
| –Ę—Ä–ł–į—Ā | 251,0 | ||
| –ü–į–Ľ–Ķ–ĺ–∑–ĺ–Ļ 291 –ľ–Ľ–Ĺ. –≥. | –ü–Ķ—Ä–ľ | 299,0 | |
| –ö–į—Ä–Ī–ĺ–Ĺ | 359,2 | ||
| –Ē–Ķ–≤–ĺ–Ĺ | 416,0 | ||
| –°–ł–Ľ—É—Ä | 443,7 | ||
| –ě—Ä–ī–ĺ–≤–ł–ļ | 488,3 | ||
| –ö–į–ľ–Ī—Ä–ł–Ļ | 542,0 | ||
| –ü—Ä–ĺ—ā–Ķ—Ä–ĺ–∑–ĺ–Ļ | |||
| –Ě–Ķ–ĺ–Ņ—Ä–ĺ—ā–Ķ—Ä–ĺ–∑–ĺ–Ļ 458 –ľ–Ľ–Ĺ. –≥. | –ē–ī–ł–į–ļ–į—Ä–ł–Ļ | 630 | |
| –ö—Ä–ł–ĺ–≥–Ķ–Ĺ | 850 | ||
| –Ę–ĺ–Ĺ–ł–Ļ | 1 000 | ||
| –ú–Ķ–∑–ĺ–Ņ—Ä–ĺ—ā–Ķ—Ä–ĺ–∑–ĺ–Ļ 600 –ľ–Ľ–Ĺ. –≥. | –°—ā–Ķ–Ĺ–ł–Ļ | 1 200 | |
| –ē–ļ—ā–į—Ā–ł–Ļ | 1 400 | ||
| –ö–į–Ľ–ł–ľ–ł–Ļ | 1 600 | ||
| –ü–į–Ľ–Ķ–ĺ–Ņ—Ä–ĺ—ā–Ķ—Ä–ĺ–∑–ĺ–Ļ 900 –ľ–Ľ–Ĺ. –≥. | –°—ā–į—ā–Ķ—Ä–ł–Ļ | 1 800 | |
| –ě—Ä–ĺ–∑–ł—Ä–ł–Ļ | 2 050 | ||
| –†—Ź—Ā–ł–Ļ | 2 300 | ||
| –°–ł–ī–Ķ—Ä–ł–Ļ | 2 500 | ||
| –ź—Ä—Ö–į–Ļ | –Ě–Ķ–ĺ–į—Ä—Ö–į–Ļ 300 –ľ–Ľ–Ĺ. –≥. | 2 800 | |
| –ú–Ķ–∑–ĺ–į—Ä—Ö–į–Ļ 400 –ľ–Ľ–Ĺ. –≥. | 3 200 | ||
| –ü–į–Ľ–Ķ–ĺ–į—Ä—Ö–į–Ļ 400 –ľ–Ľ–Ĺ. –≥. | 3 600 | ||
| –ē–ĺ–į—Ä—Ö–į–Ļ | 3 800 | ||
| –•–į–ī–Ķ–Ļ | 4 700 |
–ė–∑—ā–ĺ—á–Ĺ–ł—Ü–ł:
Hypotheses for the transition from radially symmetrical to bilaterally symmetrical animals.
Classification of the Chordates
Etienne Geoffroy Saint-Hilaire revu et corrigé par l'embryologie et la génétique -Thibaut BRUNET
The origins of vertebrate adaptive immunity
–ü–ł–ļ–į–Ļ—Ź ‚ÄĒ —Ā–į–ľ–ĺ–Ķ –Ņ—Ä–ł–ľ–ł—ā–ł–≤–Ĺ–ĺ–Ķ —Ö–ĺ—Ä–ī–ĺ–≤–ĺ–Ķ?¬†- elementy.ru
Simon Conway Morris and Jean-Bernard Caron.¬†Pikaia gracilens¬†Walcott, a stem-group chordate from the Middle Cambrian of British Columbia¬†//¬†Biological Reviews. 2012. Advance online publication (PDF, 7 –ú–Ī).
Haikouichthys - wikipedia
There’s something fishy about that fish-pigeonchess.com
Myllokunmingia- wikipedia
Haikouella - wikipedia
Pikaia gracilens, is the precursor of modern vertebrates, including humans
–ď–Ķ–Ĺ–ĺ–ľ –Ľ–į–Ĺ—Ü–Ķ—ā–Ĺ–ł–ļ–į –Ņ–ĺ–ľ–ĺ–≥ —Ä–į—Ā–ļ—Ä—č—ā—Ć —Ā–Ķ–ļ—Ä–Ķ—ā —ć–≤–ĺ–Ľ—é—Ü–ł–ĺ–Ĺ–Ĺ–ĺ–≥–ĺ —É—Ā–Ņ–Ķ—Ö–į –Ņ–ĺ–∑–≤–ĺ–Ĺ–ĺ—á–Ĺ—č—Ö-elementy.ru





















–ö–ĺ–ľ–Ķ–Ĺ—ā–į—Ä–ł
–ú–ĺ–Ľ—Ź, —Ä–Ķ–≥–ł—Ā—ā—Ä–ł—Ä–į–Ļ—ā–Ķ —Ā–Ķ –ĺ—ā T–£–ö!
–ź–ļ–ĺ –≤–Ķ—á–Ķ –ł–ľ–į—ā–Ķ —Ä–Ķ–≥–ł—Ā—ā—Ä–į—Ü–ł—Ź, –Ĺ–į—ā–ł—Ā–Ĺ–Ķ—ā–Ķ –Ę–£–ö!
24.04 2015 –≤ 11:30
–ě—Č–Ķ –Ņ–ĺ –Ĺ–Ķ–Ľ–Ķ–Ņ–ĺ –Ķ –Ķ–ī–ł–Ĺ –∑–į–≤—ä—Ä—ą–Ķ–Ĺ –≤–Ķ—á–Ķ –Ī–ĺ–∑–į–Ļ–Ĺ–ł–ļ, –ĺ—ā —ā–ł—á–į–Ĺ–Ķ –ī–į –Ľ–ĺ–≤–ł —Ä–ł–Ī–į, –ī–į —Ā–ł –∑–į–≥—É–Ī–ł –ļ—Ä–į–ļ–į—ā–į –ł –ī–į —Ā–Ķ –Ņ—Ä–Ķ–≤—ä—Ä–Ĺ–Ķ –Ņ–ĺ—Ā—ā–Ķ–Ņ–Ķ–Ĺ–Ĺ–ĺ –≤ –ļ–ł—ā! –ú–Ĺ–ĺ–≥–ĺ –Ķ –ł–Ĺ—ā–Ķ—Ä–Ķ—Ā–Ĺ–į –Ņ—Ä–Ķ—Ö–ĺ–ī–Ĺ–į—ā–į —Ą–ĺ—Ä–ľ–į, –ļ–ĺ—Ź—ā–ĺ,–ĺ—Č–Ķ –Ĺ–Ķ –ĺ—ā–ļ—Ä–ł—ā–į –ĺ—ā "—É—á–Ķ–Ĺ–ł—ā–Ķ"! –ē–í–ě–õ–ģ–¶–ė–ě–Ě–ė–°–Ę–ė-–ė-–ė-–ė!!
–ü–ĺ—Ā–Ľ–Ķ–ī–Ĺ–ł –ļ–ĺ–ľ–Ķ–Ĺ—ā–į—Ä–ł
–ü—Ä–ĺ—Ā—ā –ß–ĺ–≤–Ķ–ļ
–ö–ĺ–Ľ–ļ–ĺ –Ī—ä—Ä–∑–ĺ —Ā—ā–į–≤–į –ļ–≤–į–Ĺ—ā–ĺ–≤–ĺ—ā–ĺ –≤–Ņ–Ľ–ł—ā–į–Ĺ–Ķ? –£—á–Ķ–Ĺ–ł –≥–ĺ –ł–∑—Ā–Ľ–Ķ–ī–≤–į—ā –≤ –į—ā–ĺ—Ā–Ķ–ļ—É–Ĺ–ī–Ķ–Ĺ –ľ–į—Č–į–Ī
–ü—Ä–ĺ—Ā—ā –ß–ĺ–≤–Ķ–ļ
–ö–ĺ–Ľ–ļ–ĺ –Ī—ä—Ä–∑–ĺ —Ā—ā–į–≤–į –ļ–≤–į–Ĺ—ā–ĺ–≤–ĺ—ā–ĺ –≤–Ņ–Ľ–ł—ā–į–Ĺ–Ķ? –£—á–Ķ–Ĺ–ł –≥–ĺ –ł–∑—Ā–Ľ–Ķ–ī–≤–į—ā –≤ –į—ā–ĺ—Ā–Ķ–ļ—É–Ĺ–ī–Ķ–Ĺ –ľ–į—Č–į–Ī
–ü—Ä–ĺ—Ā—ā –ß–ĺ–≤–Ķ–ļ
–ü–ĺ—Ā–Ľ–Ķ–ī–Ĺ–į—ā–į —ā–Ķ–ĺ—Ä–Ķ–ľ–į –Ĺ–į –°—ā–ł–≤—ä–Ĺ –•–ĺ–ļ–ł–Ĺ–≥ –Ņ—Ä–Ķ–ĺ–Ī—Ä—ä—Č–į –≤—Ä–Ķ–ľ–Ķ—ā–ĺ –ł –Ņ—Ä–ł—á–ł–Ĺ–Ĺ–ĺ—Ā—ā—ā–į
–ü—Ä–ĺ—Ā—ā –ß–ĺ–≤–Ķ–ļ
–†–į–∑—Ä—Ź–∑–≤–į–Ĺ–Ķ—ā–ĺ –Ĺ–į —Ą–ĺ—ā–ĺ–Ĺ –Ĺ–į –ī–≤–Ķ —Ā—ä–∑–ī–į–≤–į –Ī–Ķ–∑–ļ—Ä–į–Ķ–Ĺ —Ä–ĺ—Ź–ļ –ĺ—ā —á–į—Ā—ā–ł—Ü–ł
zlatkov
–£—á–Ķ–Ĺ–ł —Ā–ļ–į–Ĺ–ł—Ä–į—ā 74 –ľ–ł–Ľ–ł–ĺ–Ĺ–į —Ä–į–ī–ł–ĺ—Ā–ł–≥–Ĺ–į–Ľ–į –ĺ—ā –ľ–Ķ–∂–ī—É–∑–≤–Ķ–∑–ī–Ķ–Ĺ –ĺ–Ī–Ķ–ļ—ā –∑–į –Ņ—Ä–ł–∑–Ĺ–į—Ü–ł –Ĺ–į –ł–∑–≤—ä–Ĺ–∑–Ķ–ľ–Ĺ–ł —ā–Ķ—Ö–Ĺ–ĺ–Ľ–ĺ–≥–ł–ł